











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkRevista Boliviana de Química
versión On-line ISSN 0250-5460
Rev. Bol. Quim v.37 n.5 La Paz dic. 2020
DOI: 10.34098/2078-3949.37.5.2
ARTÍCULOS ORIGINALES COMPLETOS
Measurement of morphological and biomass changes, and of the oxalic acid
contents, after IR and UV radiation in Oxalis Triangularis
Medición de los cambios morfológicos y de biomasa, y del contenido de ácido
oxálico, después de irradiación IR y UV en Oxalis Triangularis
Ruth Nina1, Carla Maldonado1, Esther Valenzuela1, Santiago Tarqui2, Armando R. Ticona-Bustillos3
Flavio Ghezzi3, Giovanna R. Almanza2,*
1 National Herbarium of Bolivia LPB, Instituto de Ecología IE, Department of Biology, Facultad de Ciencias
Puras y Naturales FCPN, Universidad Mayor de San Andrés UMSA, P.O. Box 10077, Calle Andrés
Bello s/n, Ciudad Universitaria Cota Cota, phone +59122799459, La Paz, Bolivia, www.umsa.bo
2 Laboratory of Bioorganic, Instituto de Investigaciones Químicas IIQ, Chemical Sciences Department, Facultad de Ciencias
Puras y Naturales FCPN, Universidad Mayor de San Andrés UMSA, P.O. Box 303, Calle Andrés Bello s/n, Ciudad Universitaria
Cota Cota, phone +59122792238, La Paz, Bolivia, www.iiq.umsa.bo
3 Instituto de Investigaciones Físicas IIF, Department of Physics, Facultad de Ciencias Puras y Naturales FCPN, Universidad
Mayor de San Andrés UMSA, P.O. Box 635, Calle Andrés Bello s/n, Ciudad Universitaria Cota Cota,
phone +59122792999, La Paz, Bolivia, www.umsa.bo
* Correspondent author: galmanza@fcpn.edu.bo
Received 09 16 2020 Accepted 12 17 2020 Published 12 30 2020
Abstract
In the face of climate change, the effect of solar radiation on morphology, biomass and different chemical compounds present in plants is of interest for food security. The present study seeks to contribute to knowledge of the effect of infrared (IR) and ultraviolet (UV-A, UV-B) radiation on oxalic acid concentration, biomass and morphology in Oxalis triangularis (Oxalidaceae). The species was exposed to visible light with IR, UV-A or UV-B radiation separately and a control group exposed only to visible light was established. Five weeks later, the number of leaves
was counted and morphological changes were observed. Also, íresh and dry weights oí leaves and rhizomes were obtained from each specimen, and the quantification of oxalic acid was performed using UV/Visible spectrophotometry. The results show that biomass, height and number of leaves increase with exposure to infrared radiation, while no significant differences in these parameters are found with UVA radiation. With UVB radiation, a depigmentation of the leaflet and a certain glossiness of the leaflets due to a probable increase in waxes and a decrease in petiole height were observed. With respect to oxalic acid concentration, no significant differences were observed with the different treatments, although a tendency to decrease with IR and increase with UVB was seen. These findings may have implications for the nutritional valué of plants containing oxalic acid, for this reason further studies are needed.
Keywords: Oxalic acid, Biomass, Oxalis triangularis, Infrared radiation, Ultraviolet radiation.
Resumen
El efecto que tiene la radiación solar sobre la morfología, biomasa y distintos compuestos químicos presentes en las plantas es de interés para la seguridad alimentaria frente al cambio climático. Este estudio tiene como objetivo contribuir al conocimiento del efecto de radiación infrarroja (IR) y ultravioleta (UV-A, UV-B) sobre la concentración de ácido oxálico, la biomasa y morfología en Oxalis triangularis (Oxalidaceae). Para esto, la especie fue expuesta a luz visible con radiación IR, UV-A o UV-B por separado y se estableció un grupo de control expuesto únicamente a luz visible. Cinco semanas después se contabilizó el número de hojas, se observaron los cambios morfológicos, se obtuvo el peso fresco y seco de las hojas y rizomas de cada espécimen, y la cuantificación de ácido oxálico se realizó por espectrofotometría UV/Visible. Los resultados mostraron que la biomasa, altura y número de hojas aumenta con la exposición a la radiación infrarroja, mientras que no se determinaron diferencias significativas en estos parámetros con la radiación UV-A. Con la radiación UV-B se observó despigmentación en los foliólos, un cierto brillo en los mismos por aumento probable de las ceras y una disminución en la altura del peciolo. Respecto a la concentración de ácido oxálico no se observaron diferencias significativas con los diferentes tratamientos, aunque se observa una tendencia a disminuir con IR y aumentar con UV-B. Estos hallazgos podrían tener implicaciones en plantas que contengan ácido oxálico por su valor nutricional, por esta razón se debe continuar con más estudios.
Palabras clave: Ácido oxálico, Biomasa, Oxalis triangularis, Radiación infrarroja, Radiación ultravioleta.
INTRODUCCIÓN
La radiación solar es importante para la vida de los organismos en el planeta, que por lo general es beneficiosa en el desarrollo de las plantas; sin embargo, altas intensidades pueden afectar importantes procesos en las plantas [1] como cambios en la morfología, anatomía, procesos fisiológicos y bioquímicos [2,3]. Por lo mismo, se han realizado numerosos estudios sobre los efectos de la radiación solar en las plantas y se ha demostrado que si bien hay efecto de la radiación en el crecimiento [3], también lo hay en la biosíntesis de compuestos químicos [2,4].
La radiación solar puede dividirse según la longitud de onda, y en nuestro estudio tendremos a la radiación ultravioleta (UV-B, 280-320 nmy UV-A, 320-400 nm), que tiene longitudes menores a la luz visible (400-700 nm) y la radiación infrarroja (IR, 700-1000 nm) con longitudes mayores a la visible [5]. La intensidad de radiación ultravioleta que recibe la tierra, naturalmente puede variar según el ángulo del sol, la latitud y la altitud [5]. En los últimos años, la intensidad de la radiación UV-B se incrementó en los polos de la Tierra por el adelgazamiento de la capa de ozono [6], y aumentó la temperatura por los gases del efecto invernadero [7], a consecuencia de las actividades antropogénicas.
Algunos estudios han demostrado que, como respuesta a altas radiaciones solares, las plantas producen ciertos compuestos que les sirven para minimizar los efectos negativos de las mismas [3,8]. El ácido oxálico es un ácido dicarboxílico (HOOC-COOH) que forma sales solubles e insolubles como ion oxalato [9] y cumple diferentes funciones en la planta [9-11], por ejemplo la formación de cristales de oxalato de calcio depositados sobre las hojas podría reflejar la radiación solar recibida [12]. Los oxalatos están presentes tanto en hojas, frutos, semillas o estructuras subterráneas de interés alimenticio [9]. Sin embargo, el consumo de alimentos con concentraciones elevadas de oxalatos, podría tener un efecto negativo dando lugar a cálculos renales, por la formación de cristales de oxalato de calcio. Además, en las mujeres después de la menopausia disminuye la capacidad de absorción del calcio, y el consumo de alimentos ricos en oxalato agudiza esta deficiencia [9].
Por lo mencionado, es de interés conocer si diferentes tipos de radiación podrían influir en la producción de ácido oxálico en las plantas y si esto guardaría relación con el aumento de la biomasa. Entre las principales familias de plantas que producen oxalatos está la familia Oxalidaceae [9]. Algunas de sus especies son comestibles (Ejemplo la oca Oxalis tuberosa Mol.), por lo que especies del género Oxalis resultan interesantes para su estudio en cuanto al efecto que pueda tener la radiación solar sobre su crecimiento y producción de ácido oxálico. En el presente estudio escogimos a Oxalis triangularis, una especie fácil de trabajar, pequeña e ideal para condiciones de laboratorio, ya que pueden crecer tanto con poca luz como a plena luz solar. En base a la información expuesta, este estudio pretende determinar el efecto de la radiación UV-A, UV-B e IR, sobre la concentración de ácido oxálico, biomasa y características morfológicas en Oxalis triangularis.
EXPERIMENTAL
Descripción de la especie de estudio
Oxalis, un género de la familia Oxilidaceae, posee una distribución cosmopolita con más de 800 especies, tiene un amplio rango altitudinal de distribución y se encuentra en diferentes climas. Presenta órganos subterráneos que le permiten pasar la estación desfavorable bajo el suelo en forma de tubérculos, rizomas o bulbos. En Bolivia se tiene el registro de 57 especies de las cuales 12 son endémicas y entre las especies alimenticias la más conocida esta Oxalis tuberosa por sus tubérculos [15,16].
En este estudio se trabajó con Oxalis triangularis, poco conocida como comestible [15] y son más utilizadas en la jardinería. Está especie tiene rizomas anaranjados, hojas compuestas trifoliadas, con foliólos triangulares de color púrpura oscuro y peciolos de 12-20 cm. La inflorescencia está dispuesta en cimas, las flores son de color rosado, con pedúnculos de similar tamaño de los peciolos (Figura 1). Es originaria de Bolivia, Brasil, Paraguay y el norte de Argentina [16].

Obtención del material vegetal
En el vivero del Jardín Botánico La Paz, ubicado en el Campus Universitario de Cota Cota, de la Universidad Mayor de San Andrés (16°30'S, 68°7'O), se sembraron aproximadamente 20 plantines de esta especie con rizomas, que se obtuvieron de un vivero comercial de Cota Cota. A partir de éstos se obtuvieron más rizomas en el transcurso de un año (aproximadamente entre 6 a 16 rizomas por plantín). Éstos fueron puestos en macetas que contenían turba, abono y tierra negra (3:1:1) y la frecuencia de riego fue de dos veces por semana. Al finalizar este periodo el total de rizomas que se obtuvieron, y con los que empezamos el trabajo, fue de 128.
Tratamientos de experimentación
Los 128 rizomas se dividieron en cuatro grupos de 32. El primero fue tratado como el grupo control, sometido durante toda la experimentación a la radiación de 8 lámparas fluorescentes que emiten luz visible (Philips TL-D 36W/54-765) por 12 horas/día. Los otros tres grupos de individuos tuvieron las mismas condiciones que el grupo control, incluida la radiación visible, pero además se adicionaron tres distintos tipos de radiaciones a cada uno:
- TRATAMIENTO 1: IR, dos lámparas incandescentes infrarrojas (Alie 250 W, E27) 6 horas/día.
- TRATAMIENTO 2: UV-A, cuatro lámparas UVA (F40T12 40 W) 8 horas/día.
- TRATAMIENTO 3: UV-B, una lámpara UVB (FS40T12-UVB-BP) lhora/día, descontinuada es decir 15 minutos por cada dos horas.
La temperatura, humedad relativa, PAR (radiación fotosintéticamente activa), intensidad de radiación UV-A, UV-B, así como el tiempo de irradiación a los que fueron sometidos cada uno de los tratamientos se presenta en la Tabla 1. Todas las lámparas estaban dispuestas aproximadamente a 50 cm de altura sobre los plantines.
Después de dos semanas de irradiación, se observó que no todos los plantines habían brotado, por lo que, a fin de contar con individuos expuestos por un mismo periodo de tiempo a las distintas radiaciones, se eliminaron de la experimentación aquellas que no habían brotado, lo que nos dejó con un número reducido de individuos por tratamiento: 19 para IR, 15 para UV-A, 18 para UV-B y 19 para el control. Estos individuos quedaron entonces expuestos por un total de cinco semanas de irradiación.

Toma de datos morfológicos
Al finalizar las cinco semanas de irradiación se tomaron los datos morfológicos. Se anotaron las características morfológicas en cada caso, como ser color, brillo y rigidez de la hoja, y luego se procedió a contar el número de hojas presentes. Finalmente, se colectaron las hojas (foliólos y peciolos por separado) y rizomas para pesarlos y así obtener el peso fresco y seco de cada individuo. Para obtener el peso seco, se dejó a temperatura ambiente en oscuridad aproximadamente dos meses y los rizomas que continuaban húmedos se los puso en un liofilizador.
Análisis de ácido oxálico
- Extracción
Para esta fase, se consideraron a los individuos que alcanzaron el peso fresco de 1 g de foliólos por planta. Como en cada tratamiento quedó un número distinto de individuos, se decidió trabajar en todos los casos con 11 individuos de Oxalis triangularis por cada tratamiento. La extracción se realizó mediante la metodología de Presswood et al. [17]. Se utilizó lg fresco de foliólos que posteriormente se secaron, estos fueron picados aproximadamente 1 mm y puesto en un matraz con 20 mi de HC1 0.2 M. La mezcla se llevó a baño María a 80 °C por 20 min, luego se dejó enfriar a temperatura ambiente por 10 min. Una vez frío se filtró y se aforó a 25 mi con HC10.2 M. Esta solución se centrifugó a 2800 rpm por 15 min (Centrífuga Macro Presvac DCS-16 RV), y se conservó el sobrenadante de la extracción.
- Extracción en fase sólida (SPE)
Para eliminar antocianinas y fenólicos del sobrenadante de la extracción, se realizó extracción en fase sólida. Se utilizaron cartuchos con 100 mg de Silicagel C18 (Discovery DSC-18 SPE 100 mg), acondicionados con 0.4 mi de metanol y 0.6 mi de agua desionizada. Posteriormente se hizo pasar por los cartuchos 1.4 mi del sobrenadante de la extracción, recuperándose una solución transparente que se usó para realizar la cuantificación de oxalatos por espectrofotometría visible. Este procedimiento se realizó para cada muestra por triplicado.
- Espectrofotometría visible
Las mediciones de ácido oxálico por espectrofotometría visible (Espectrofotómetro SPEXTRO QUANT Pharo 300) se realizaron utilizando 1 mi de solución de la muestra obtenida por SPE por triplicado de cada planta, mediante el método de Zhai et al. [18]. En un vaso de precipitado de 50 mi, se vertió 2 mi H2SO4 0.1 M, y 0.8 mi de RhB (rodamina B) 1.75 x 10-5 M. Después se agregó lml de la solución obtenida de la muestra por SPE y 0.4 mi de K2Cr2O7 3.0x 1O-5 M, esta mezcla se calentó a 50 °C durante 10 min. Luego se dejó enfriar a temperatura ambiente por 10 minutos. Finalmente, se agregó 0.8 mi de NaOH 4.0 M y se trasvasó la solución a un matraz aforado de 10 mi, aforando con agua des ionizada.
La determinación de la concentración de ácido oxálico se realizó a 552 nm, con la curva de calibración previamente obtenida con la metodología de Zhai et al. [18] (Figura 2).

Análisis estadístico
Para analizar el efecto de la radiación en la biomasa y concentración de ácido oxálico en Oxalis triangularis se revisó la normalidad de los datos con la prueba de Shapiro-Wilk y homogeneidad de varianzas con la prueba de Levene, y se escogió utilizar la prueba de Kruskal-Wallis para muestras no paramétricas, seguida de una comparación entre pares, mediante la corrección de Bonferroni con un nivel de significancia de 0.05 %. La prueba estadística y gráficos se realizaron con el software RStudio [19].
RESULTADOS Y DISCUSIÓN
Características morfológicas
Después de las cinco semanas de exposición, los individuos de Oxalis triangularis mostraron las siguientes características que se detallan en la Tabla 2 y la Figura 3.
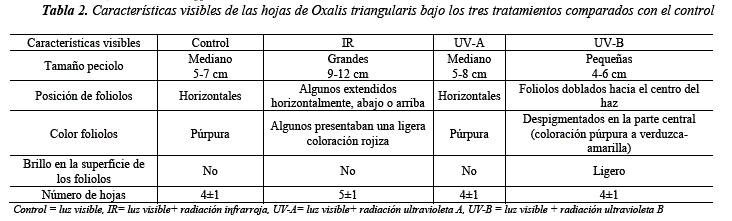

La coloración rojiza que presentaron los individuos en el tratamiento con radiación IR (Tabla 2, Figura 3B), podría deberse a niveles de FR (rojo lejano). Esta suposición se basa en otros estudios que probaron que ante FR, la concentración tanto de clorofila como de antocianinas tienden a disminuir [20]. El grupo control en este estudio muestra que las hojas tienen una coloración púrpura debida a la presencia de antocianinas en sus hojas [17,23], por lo que, la perdida de coloración púrpura de los foliólos de Oxalis triangularis pudo haber ocurrido por la disminución de esos pigmentos, ya que este tratamiento tiene lámparas incandescentes infrarrojas que emiten principalmente en FR.
Otro carácter morfológico de Oxalis triangularis que cambió notablemente con la radiación IR fue la altura de los peciolos (Tabla 2, Figura 3B). Estudios en las especies como Stellaria longipes y Arabidopsis thaliana, muestran tallos más grandes con niveles bajos del cociente R/FR (rojo/rojo lejano) [24,25]. Sin embargo, cuando crecen solo bajo radiación infrarroja son más pequeñas como en Avena sativa [24]. En el tratamiento IR, como se mencionó, tenía lámparas infrarrojas más luz visible, y cuando hay niveles bajos de R/FR se alargan los tallos para evitar la sombra por otras plantas que es percibida por la calidad de la luz [27,28]. Por lo tanto, el aumento de crecimiento de peciolos pudo ser respuesta para alcanzar luz.
Los individuos expuestos al tratamiento UV-A, no mostraron diferencias en tamaño o características morfológicas respecto al Control (Tabla 2, Figura 3C). Con Betula péndula encontraron que ni la radiación UV-A ni la UV-B tenían efecto sobre la altura de la planta [29,30], sin embargo, otra investigación con Mentha piperita mostró que el tamaño de las hojas podría aumentar con la radiación UV-A [29]. En Lactuca sativa aumentó el tamaño de las hojas además del peso seco con UV-A a 402 nm más luz roja y azul, pero ambos caracteres disminuyeron a 387 y 367 nm más luz roja y azul [30]. Es probable que la UV-A tenga un efecto diferente en cada especie y según la longitud de onda del rango UV-A, siendo que en algunas induce al crecimiento foliar y en otros no. Las lámparas UV-A que tuvimos en este estudio presentan el pico emisión a 350 nm, y no se observó ningún efecto notable en la especie de estudio.
En el tratamiento con UV-B, los foliólos de Oxalis triangularis se curvaron hacia el haz y se mostraron más brillosos que en los otros tratamientos (Figura 3D), esta sería una respuesta fotomorfogénica para disminuir el área expuesta a los rayos ultravioleta [31] y un incremento de ceras en la cutícula para refractar la radiación ultravioleta [32] como se obtuvo en hojas de Lactuca sativa [33]. Asimismo, los foliólos de Oxalis triangularis presentaban una despigmentación, esto podría ser por las quemaduras de la radiación, a consecuencia de la inhibición acumulada de clorofilas y antocianinas en los foliólos [34]. Por lo que, estos cambios observados serían producto de la UV-B, ya que, en el Control, donde la exposición fue únicamente a luz visible, las hojas son púrpura por las antocianinas presentes.
Por otro lado, el largo de los peciolos fue menor en el tratamiento UV-B (Tabla 2, Figura 3D). Siendo que la UV-B puede interferir con el metabolismo del ácido indolacético, que a su vez inhibiría el crecimiento de las plantas [35], este es uno de los efectos documentados que se observa cuando se induce a las plantas a UV-B [31] y en comparación del Control, este grupo de individuos presentaron el menor crecimiento.
Biomasa
La biomasa de rizomas para los diferentes tratamientos no fue significativamente diferente respecto al Control (x2 =2.31, gl=3, p=0.51). En el tratamiento IR, aumentó la biomasa de foliólos (x2 =31.83, gl=3, p<0.05) y peciolos (x2 =45.57, gl=3, p<0.05). En los tratamientos UV-A y UV-B, no se observaron diferencias en cuanto a la biomasa respecto el Control (Figura 4 D, E y F). Sin embargo, se observa una tendencia de disminución de la biomasa de foliólos y peciolos en el tratamiento UV-B.

El aumento de la biomasa de foliólos y peciolos de los individuos bajo el tratamiento IR, estaría directamente relacionado con el tamaño y el aumento en el número de hojas (Figura 4B, 6). En Lactuca sativa se obtuvo que luz visible más FR aumenta la altura del tallos, tamaño de las hojas y la biomasa seca y fresca [20]. Esta podría ser respuesta a niveles bajos de R/FR dado que este valor se constituye en una señal de advertencia de la presencia de vegetación competidora estimulando el crecimiento de las plantas para alcanzar fuentes de luz [36]. En Stellaria longipes, aumenta de tamaño del tallo, debido a la regulación de genes que llevan a la expansión de la pared celular cuando los niveles de R/FR son bajos [23], es probable que Oxalis triangularis al aumentar de tamaño también incremento en biomasa por las condiciones del tratamiento. Por otro lado, en los tratamientos UV-A y UV-B, la biomasa de rizomas, foliólos y peciolos no mostró diferencia estadísticamente significativa con el Control (Figura 4). Similar resultado se obtuvo en Betulapéndula [29,30].
Concentración de ácido oxálico
En la figura 5 se observa que, aunque hay una ligera tendencia de incremento de ácido oxálico con el tratamiento UV-B, una disminución con el tratamiento IR, y ninguna reacción del tratamiento UV-A comparado con el Control, estadísticamente no hay una diferencia significativa entre tratamientos (x2 =7.12, gl=3, p=0.07).
Un análisis de componentes principales (figura 6) confirma que las variables de biomasa están positivamente correlacionadas con la exposición a radiación IR. Sin embargo, la producción de ácido oxálico parece tener más relación con la exposición a las radiaciones UV-B, seguida de UV-A.


La inexistencia de una diferencia significativa en producción de ácido oxálico entre tratamientos podría deberse al tiempo de crecimiento de la especie en estudio. Estudios con Capsicum annuum (hierba anual de hasta un metro de alto), mostraron cristales de oxalato de calcio casi ausentes en plantas con 3 semanas de crecimiento [37] que crecieron con bajos niveles FR/R y con Beta vulgaricus (hierba de hasta 2 m de alto) se vio un incremento en la concentración de ácido oxálico con la radiación UV-B después de 8 semanas de crecimiento [17]. Estas especies, si bien tienen una rapidez de crecimiento distinto, muestran también producción de ácido oxálico en diferentes etapas. En nuestro caso, Oxalis triangularis es una hierba que no sobrepasa los 30 cm de altura y se esperó que la planta llegue a su producción máxima después de 5 semanas, sin embargo, puede ser que con un tiempo más de exposición, hubiésemos tenido otros resultados. Aunque, en otro estudio menciona que los cultivares con rápido crecimiento de Spinacia olerácea (hierba de hasta 1 m de altura), disminuye el contenido de ácido oxálico en hojas muestreadas después de casi 7 semanas [38], talvez el análisis de ácido oxálico podría depender de la especie de estudio y en este caso, valdría la pena hacer un seguimiento de los niveles de concentración de ácido oxálico en varias etapas de crecimiento.
En nuestro estudio, la respuesta de ácido oxálico que tuvo la UV-A fue menos evidente (Figura 6). El efecto que puede producir la UV-A es menor que el que se tiene con la UV-B en el sistema fotosintético [39], tal vez la producción de ácido oxálico no se ve influenciada por longitudes de onda menos energéticas como es la UV-A a comparación de la UV-B. Además que la UV-A puede activar la expresión de un conjunto de genes distintos a los que produce la UV-B [39].
Se conoce que existen moléculas precursoras de otras y en estudios se ha comprobado que la UV-B puede incrementar la concentración de moléculas como el ácido ascórbico [40,41] este es el principal precursor del ácido oxálico [9,42], y se podría pensar que debido a esto el ácido oxálico se incrementaría con UV-B. Aunque, otros estudios muestran que en Spinacia olerácea [43] y el Hibiscus sabdariffa, no hubo relación entre ambas moléculas, ya que fue mayor el contenido de ácido ascórbico y menor el de ácido oxálico en hojas que crecieron con mayor intensidad de luz [44].
CONCLUSIONES
Nuestros principales resultados mostraron que, de los tres tipos de radiación utilizados, sólo la luz IR tiene efecto positivo sobre el aumento de la biomasa en Oxalis triangularis, éste aumento fue de casi 120% en hojas con relación a nuestro control, 70% en el peciolo y 25% en número de hojas. Sin embargo, no encontramos una relación clara de la producción de ácido oxálico en ninguno de los casos, aunque si una ligera tendencia de aumento con UV-B y disminución con IR. Estas tendencias, coinciden con otros estudios para otras especies [17,37], por lo que valdría la pena analizar otras variables o ampliar el estudio tomando en cuenta por ejemplo más tiempo de exposición a diferentes radiaciones o una combinación de ellas.
RECONOCIMIENTOS
Este trabajo se realizó gracias a una beca "Erika Geyger" del Instituto de Ecología a Ruth Nina. Agradecemos también al Dr. Carlos Molina por el apoyo en el análisis estadístico y a Magaly Mercado por las sugerencias al manuscrito.
REFERENCIAS
1. Carrasco-Ríos, L. 2009, Efecto de la radiación ultravioleta-B en plantas, Idesia (Arica), 27(3), 59-76. DOI: 10.4067/S0718-34292009000300009 [ Links ]
2. Kataria, S., Baroniya, S. S., Kanungo M., Bhaghel, L. 2014, Effect of Exclusion of Solar UV radiation on Plants, Plant Science Today, 1(4), 224-232. DOI: 10.14719/pst.2014.1.4.61 [ Links ]
3. Neugart, S., Schreiner, M. 2018, UVB and UVA as eustressors in horticultural and agricultural crops. Scientia Horticulturae 234, 370-381. DOI: 10.1016/j.scienta.2018.02.021 [ Links ]
4. Zhang, W. J., Björn, L. O. 2009, The effect of ultraviolet radiation on the accumulation of medicinal compounds in plants, Fitoterapia, 80(4), 207-218.
5. Madronich, S., McKenzie, R. L., Björn, L. O., Caldwell M. M. 1998, Changes in biologically active ultraviolet radiation reaching the Earth's surface. Journal of Photochemistry and Photobiology B: Biology 46(1-3), 5-19. DOI: 10.1016/S1011-1344(98)00182-1
6. Wachowski, L., Kirszensztejn P., Foltynowicz, Z. 2001, Ecological replacements of ozone-depleting substances, Polish Journal of Environmental Studies, 10(6), 415-435 [ Links ]
7. Olivo, M. D. L., Soto-Olivo, A. 2010, Comportamiento de los gases de efecto invernadero y las temperaturas atmosféricas con sus escenarios de incremento potencial, Universidad, Ciencia y tecnología, 14(57), 221-230. [ Links ]
8. Bian, Z. H., Yang, Q. C., Liu, W. K. 2015, Effects of light quality on the accumulation of phytochemicals in vegetables produced in controlled environments: a review. Journal of the Science of Food and Agriculture, 95(5), 869-877. DOI: 10.1002/jsfa.6789 [ Links ]
9. Noonan S. N., Savage G. S. 1999, Oxalate content of foods and its effect on humans, Asia Pacific journal of clinical nutrition, 8(1), 64-74. [ Links ]
10. Korth, K. L., Doege, S. J., Park, S. H., Goggin, F. L., Wang, Q., Gomez, S. K., Nakata, P. A. 2006, Medicago truncatula mutants demonstrate the role of plant calcium oxalate crystals as an effective defense against chewing insects, Plant Physiology, 141(1), 188-195. DOI:10.1104/pp.106.076737 [ Links ]
11. Weir, T. L., Bais, H. P., Stull, V. J., Callaway, R. M., Thelen, G. C., Ridenour, W. M., Vivanco, J. M. 2006, Oxalate contributes to the resistance of Gaillardia grandiflora and Lupinus sericeus to a phytotoxin produced by Centaurea maculosa, Planta, 223(4), 785–795. DOI: 10.1007/s00425-005-0192-x
12. Jacobsen, S. E., Mujica, A., Jensen, C. R. 2003, The resistance of quinoa (Chenopodium quinoa Willd.) to adverse abiotic factors. Food Reviews International, 19(1-2), 99-109. DOI: 10.1081/FRI-120018872 [ Links ]
13. Oberlander, K. C., Emshwiller, E., Bellstedt, D. U., Dreyer, L. L. 2009, A model of bulb evolution in the eudicot genus Oxalis (Oxalidaceae), Molecular Phylogenetics and Evolution, 51(1), 54-63. DOI: 10.1016/j.ympev.2008.11.022 [ Links ]
14. Jørgensen P. M., Nee M.H., Beck S.G. 2014, Catálogo de las plantas vasculares de Bolivia, Monographs in Systematic Botany from the Missouri Botanical Garden, 127(1–2), 1744.
15. Pazmiño-Durán, E. A., Giusti, M. M., Wrolstad, R. E., Glória, M. B. A. 2001, Anthocyanins from Oxalis triangularis as potential food colorants, Food Chemistry, 75(2), 211-216. DOI: 10.1016/S0308-8146(01)00201-1 [ Links ]
16. Lourteig, A. 2000, Oxalis L. Subgenero Monoxalis (Small) Lourt., Oxalis y Trifidus Lourt. Bradea, 7 (2):629. [ Links ]
17. Presswood, H., Hofmann, R., Savage, G. P. 2012, Effects of UV-B radiation on oxalate content of silver beet leaves, Journal of Food Research, 1(4),1–6. DOI: 10.5539/jfr.v1n4p1
18. Zhai, Q. Z., Zhang, X. X., Liu, Q. Z. 2006, Catalytic kinetic spectrophotometry for the determination of trace amount of oxalic acid in biological samples with oxalic acid–rhodamine B–potassium dichromate system, Spectrochimica Acta Part A: Molecular and Biomolecular Spectroscopy, 65(1), 1-4. DOI: 10.1016/j.saa.2005.07.080
19. RStudio Team. 2020, RStudio: Integrated Development for R. RStudio, PBC, Boston, MA URL http://www.rstudio.com/. [ Links ]
20. Li, Q., Kubota, C. 2009, Effects of supplemental light quality on growth and phytochemicals of baby leaf lettuce, Environmental and Experimental Botany, 67(1), 59-64. DOI: 10.1016/j.envexpbot.2009.06.011 [ Links ]
21. Nielsen, S. L., Simonsen, A. M. 2011, Photosynthesis and photoinhibition in two differently coloured varieties of Oxalis triangularis-the effect of anthocyanin content, Photosynthetica, 49(3), 346-352. DOI: 10.1007/s11099-011-0042-y [ Links ]
22. Alokam, S., Chinnappa, C. C., Reid D. M. 2002, Red/far-red light mediated stem elongation and anthocyanin accumulation in Stellaria longipes: differential response of alpine and prairie ecotypes, Canadian Journal of Botany, 80(1): 72-81. DOI: 10.1139/b01-137 [ Links ]
23. Sasidharan, R., Chinnappa, C. C., Voesenek, L. A., Pierik, R. 2008, The regulation of cell wall extensibility during shade avoidance: a study using two contrasting ecotypes of Stellaria longipes, Plant Physiology, 148(3), 1557-1569. DOI: 10.1104/pp.108.125518 [ Links ]
24. Johnson, C. F., Brown, C. S., Wheeler, R. M., Sager, J. C., Chapman, D. K., Deitzer, G. F. 1996, Infrared light‐emitting diode radiation causes gravitropic and morphological effects in dark‐grown oat seedlings, Photochemistry and photobiology, 63(2), 238-242. DOI: 10.1111/j.1751-1097.1996.tb03020.x [ Links ]
25. Gommers, C. M., Visser, E. J., St Onge, K. R., Voesenek, L. A., Pierik, R. 2013, Shade tolerance: when growing tall is not an option, Trends in plant science, 18(2), 65-71. DOI: 10.1016/j.tplants.2012.09.008 [ Links ]
26. Carvalho, R. F., Takaki, M., Azevedo, R. A. 2011, Plant pigments: the many faces of light perception, Acta Physiologiae Plantarum, 33(2), 241-248. [ Links ]
27. Kotilainen, T., Venäläinen, T., Tegelberg, R., Lindfors, A., Julkunen‐Tiitto, R., Sutinen, S., Aphalo, P. J. 2009, Assessment of UV biological spectral weighting functions for phenolic metabolites and growth responses in silver birch seedlings, Photochemistry and Photobiology, 85(6), 1346-1355. DOI: 10.1111/j.1751-1097.2009.00597.x [ Links ]
28. Morales, L. O., Tegelberg, R., Brosché, M., Keinänen, M., Lindfors, A., Aphalo, P. J. 2010, Effects of solar UV-A and UV-B radiation on gene expression and phenolic accumulation in Betula pendula leaves, Tree Physiology, 30(7), 923-934. DOI: 10.1093/treephys/tpq051 [ Links ]
29. Maffei, M., Canova, D., Bertea, C. M., & Scannerini, S. 1999, UV-A effects on photomorphogenesis and essential-oil composition in Mentha piperita, Journal of Photochemistry and Photobiology B: Biology, 52(1-3), 105-110. DOI: 10.1016/S1011-1344(99)00110-4 [ Links ]
30. Samuolienė, G., Viršilė, A., Miliauskienė, J., Haimi, P., Laužikė, K., Jankauskienė, J., Brazaitytė, A. 2020, The Photosynthetic Performance of Red Leaf Lettuce under UV-A Irradiation, Agronomy, 10(6), 761.
31. Jansen, M. A., Gaba, V., Greenberg, B. M. 1998, Higher plants and UV-B radiation: balancing damage, repair and acclimation, Trends in plant science, 3(4), 131-135. [ Links ]
32. Holmes, M. G. Keiller D. R. 2002, Effects of pubescence and waxes on the reflectance of leaves in the ultraviolet and photosynthetic wavebands: a comparison of a range of species, Plant, Cell & Environment, 25(1), 85-93. DOI: 10.1046/j.1365-3040.2002.00779.x [ Links ]
33. Lee, M. J., Son, J. E., Oh, M. M. 2014, Growth and phenolic compounds of Lactuca sativa L. grown in a closed‐type plant production system with UV‐A, ‐B, or‐C lamp, Journal of the Science of Food and Agriculture, 94(2), 197-204. DOI: 10.1002/jsfa.6227 [ Links ]
34. Liu, Y., Chen, N., Zuo, C., Wu, Y., Che, F., Chen, B. 2019, The mechanism of color fading in sunburned apple peel, Acta Physiologiae Plantarum, 41(1), 2. DOI: 10.1007/s11738-018-2792-7 [ Links ]
35. Ros, J., Tevini, M. 1995, Interaction of UV-radiation and IAA during growth of seedlings and hypocotyl segments of sunflower, Journal of Plant Physiology, 146(3), 295-302. [ Links ]
36. Franklin, K. A. 2008, Shade avoidance, New Phytol, 179, 930–944. DOI: 10.1111/j.1469-8137.2008.02507.x
37. Schuerger, A. C., Brown, C. S., Stryjewski, E. C. 1997, Anatomical features of pepper plants (Capsicum annuum L.) grown under red light-emitting diodes supplemented with blue or far-red light, Annals of Botany, 79(3), 273-282. DOI: 10.1006/anbo.1996.0341 [ Links ]
38. Hirooka M, Sugiyama N. 1992, Effect of growth rates on oxalate concentrations in spinach leaves, Journal of the Japanese Society for Horticultural Science, 61, 575–579. DOI: 10.2503/jjshs.61.575
39. Zhou, B., Li, Y., Xu, Z., Yan, H., Homma, S., Kawabata, S. 2007, Ultraviolet A-specific induction of anthocyanin biosynthesis in the swollen hypocotyls of turnip (Brassica rapa), Journal of Experimental Botany, 58(7), 1771-1781. DOI: 10.1093/jxb/erm036 [ Links ]
40. Majer, P., Hideg, É. 2012, Existing antioxidant levels are more important in acclimation to supplemental UV-B irradiation than inducible ones: studies with high light pretreated tobacco leaves, Emirates Journal of Food and Agriculture, 24(6), 598-606. DOI: 10.9755/ejfa.v24i6.14678 [ Links ]
41. Sakalauskaite, J., Viškelis, P., Duchovskis, P., Dambrauskiene, E., Sakalauskiene, S., Samuoliene, G., & Brazaityte, A. 2012, Supplementary UV-B irradiation effects on basil (Ocimum basilicum L.) growth and phytochemical properties. Journal of Food, Agriculture & Environment, 10(3&4), 342-346.
42. Kostman, T. A., Tarlyn, N. M., Loewus, F. A., Franceschi, V. R. 2001, Biosynthesis of L-ascorbic acid and conversion of carbons 1 and 2 of L-ascorbic acid to oxalic acid occurs within individual calcium oxalate crystal idioblasts. Plant Physiology, 125(2), 634-640. DOI: 10.1104/pp.125.2.634 [ Links ]
43. Proietti, S., Moscatello, S., Colla, G., Battistelli, Y. 2004, The effect of growing spinach (Spinacia oleracea L.) at two light intensities on the amounts of oxalate, ascorbate and nitrate in their leaves, The Journal of Horticultural Science and Biotechnology, 79(4), 606-609. DOI: 10.1080/14620316.2004.11511814 [ Links ]
44. Mataa, M., Makungu, B., Siziya, I. 2018, Shading effects of intercropping roselle (Hibiscus sabdariffa) genotypes on plant development, assimilate partitioning and leaf nutrient content, International Journal of Agricultural Research, Innovation and Technology, 8(1), 7-13. DOI: 10.3329/ijarit.v8i1.38223 [ Links ]

