











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkIntroducción
Bolivia es un país megadiverso, ya sea desde una perspectiva botánica como faunística Ibisch 1998. Posee además una gran diversidad de biomas o ecorregiones (Ribera et al. 1997, Navarro 2002, Josse 2003. Esta gran biodiversidad se puede explicar por el tamaño del país (más de un 1 000 000 km2), lo que implica una gran extensión latitudinal y longitudinal, y por tanto la existencia de un amplio gradiente ambiental. La importante extensión longitudinal hace que Bolivia posea tierras bajas y además regiones de alta montaña, ya que los Andes se encuentran incluidos en su territorio en su cuadrante suroeste. Los Andes generan gran heterogeneidad espacial, lo que también ayuda a explicar la alta diversidad del país. Lamentablemente, el conocimiento de la biota de Bolivia es uno de los menos desarrollados del continente, entre otras cosas porque en el país se empezó a hacer biología recién desde los años 70.
Gran parte de los esfuerzos científicos se ha dirigido al conocimiento del norte del país, que es donde se encuentra la región amazónica en todas sus variantes. De la parte sur, más árida y estacional, se conoce realmente muy poco. Esto es lamentable dado que en la mitad sur se encuentran varias de las ecorregiones del país (Ribera et al. 2006, Navarro 2010), las cuales incrementan su diversidad más allá de lo estrictamente amazónico. Por otro lado, a pesar de su escasez general, la mayor parte del conocimiento se traduce en listas de flora y fauna. Si hablamos de aspectos de ecología o genética de estas especies y de las comunidades que estas forman, el conocimiento es casi nulo. Por ejemplo, ¿de cuántas de las ≈ 15.000 especies de plantas vasculares que existen en el país conocemos la autoecología?
Este desconocimiento es hoy tanto más importante cuanto que nos encontramos en un momento de cambio climático, y considerando que hoy posiblemente somos más conscientes de lo que implica la conservación de las áreas naturales en términos de servicios ecosistémicos. De igual manera, el no entender las peculiaridades de nuestros sistemas ecológicos es malo porque en ese conocimiento se sustenta cualquier intento serio encaminado a la conservación, manejo y restauración de comunidades biológicas y especies Larrea-Alcázar et al. 2013. Por si fuera poco, en los últimos años se ha producido una importante degradación de las masas boscosas del país Colque 2022. Estas son algunas de las razones que hacen que sea imperativo seguir realizando estudios de ecología fundamental de especies, poblaciones y comunidades de organismos presentes en el país. Y en este marco se inserta el presente estudio, que trata de brindar información sobre una especie endémica de Bolivia, y específicamente endémica de una de las ecorregiones del país: la Prepuna boliviana.
Bulnesia rivas-martinezii (Bulnesia, de aquí en adelante; Zygophyllaceae) es un endemismo del sur de Bolivia Navarro 1994. Se encuentra en la Prepuna boliviana, aunque existe una población disyunta en la zona más árida de los valles secos andinos (Oronkhota). La Prepuna boliviana es una de las ecorregiones bolivianas que entra en la categoría de desierto (semidesierto, más exactamente, ver López 2000), y fue explícitamente descrita hace más de 20 años López 2000,López & Beck 2002, aunque ya en 1994 Navarro mencionaba una “Puna o Prepuna xerofítica de Cabrera & Willink”, haciendo alusión a esta ecorregión Navarro 1994. Bulnesia es un arbusto bajo, siempreverde y con copas muy extendidas horizontalmente, cuya distribución en la Prepuna se da exclusivamente en un sector específico: el valle de los Cintis y algunas áreas vecinas, entre 2.300-3.150 m (obs. pers,).
Bulnesia es un género de los desiertos sudamericanos Palacios & Hunziker 1984. En la Prepuna argentina se encuentra Bulnesia schickendantzii, la cual se supondría es una especie vicaria de B. rivas-martinezii. Sin embargo, esta última está más emparentada con Pintoa chilensisBohnert et al. 2020, que crece en un pequeño sector del desierto de Atacama (Región de Atacama, Chile). Esto es otro ejemplo de lo intrincado de la biogeografía sudamericana, en la cual el levantamiento de los Andes fue uno de los eventos más significativos. Es también un claro ejemplo de que la taxonomía del “clado-Larrea” Bohnert et al. 2020 debe todavía trabajarse.
Dado que es fundamental contar con datos de campo que nos permitan conocer la biología de las especies, en especial si las especies tienen áreas de distribución muy restringidas que las hagan susceptibles de extinción, en este artículo principalmente hago un análisis de los patrones espaciales de Bulnesia en diferentes lugares de su área de distribución. El estudio de los patrones espaciales, gracias al uso de una variedad de modelos nulos, permite inferir la existencia de los procesos ecológicos (facilitación, competencia, mortalidad) que pueden estar actuando en una comunidad biológica (e.g., Wiegand & Moloney 2004,2014). También describo la abundancia de las cuatro poblaciones estudiadas (y de las especies acompañantes), y la abundancia por clases de tamaño (altura/ancho de copa). Creo que este tipo de información puede ser muy útil para, eventualmente, realizar un plan de manejo de la especie, la cual, debido a su peculiaridad biogeográfica, se constituye en una especie de alto valor biológico para Bolivia.
Métodos
El estudio está basado principalmente en datos obtenidos en un trabajo de campo realizado en 2017 en el cual se mapearon las coordenadas cartesianas de todos los individuos de leñosas ubicados dentro de parcelas distribuidas en diferentes localidades de la Prepuna boliviana (ver López et al. 2020, para ulteriores detalles) y en observaciones personales que vengo realizando desde comienzos de los años 2000 en la Prepuna, tanto la boliviana como la argentina. También incluyo alguna información a partir de un estudio realizado en 2006, en el que estudié la distribución de plántulas de leñosas y cactáceas y determiné las coberturas de las especies de arbustos en 20 localidades de la Prepuna bolivianoargentina.
En 2017, junto a un equipo de biólogos realicé un estudio dirigido a determinar los patrones espaciales de las especies leñosas de la Prepuna boliviana. Se muestrearon nueve localidades. En tres de estas localidades encontramos poblaciones de Bulnesia: Cruzhuasa (20,754S, 65,234W, 2.400 m), Cruzhuasa 2 (20.837S, 65.227W, 2.400 m) y Culpina (20.735S, 65.205W, 2.700 m). En cada una de las localidades se establecieron parcelas (de tamaño variable según las características del lugar; ver Tabla 1). Las parcelas se establecieron en lugares con uniformidad en términos de características topográficas y vegetacionales (misma exposición y pendiente, misma formación vegetal, etc.), requisito importante para interpretar los patrones espaciales en términos de interacciones entre las plantas. Los lugares elegidos quedan más o menos lejos de centros poblados, y se encontraban en aparente buen estado de conservación en el momento de los estudios. Sin embargo, en casi toda la región de la Prepuna hay actividad ganadera relacionada principalmente con cabras (Capra aegragus hircus), que seguramente tienen efecto sobre el crecimiento de los arbustos palatables y el reclutamiento de nuevos individuos (por herbivoría o simple pisoteo). En los resultados se incluyeron los datos tomados en la localidad de Alto Impora (21,450S, 65,559W, 3.000 m) en 2006, donde no se realizó el mapeo de los individuos, pero sí se seleccionaron arbustos al azar para otro estudio que se realizó entonces. Para todos los arbustos de las cuatro localidades se midieron la altura y el tamaño de copa. Por otro lado, también se midió la cobertura de arbustos mediante la línea de intercepción, la cual permite determinar simultáneamente la cobertura absoluta y la relativa de la vegetación leñosa, que es lo que se reporta en este estudio. La cobertura absoluta nos dice cuánto de una determinada superficie está cubierto por las copas de los arbustos, mientras que la cobertura relativa nos informa sobre el porcentaje del total de cobertura vegetal que corresponde a Bulnesia. La línea de intercepción es un mejor medio para determinar la cobertura absoluta que la suma de los tamaños de las copas de los arbustos, pues frecuentemente las comunidades presentan estratificación (algunos individuos crecen debajo de arbustos mayores) y eso hace que se sobreestime la cobertura real Mueller- Dombois & Ellenberg 1974,Kent & Coker 1992. Las cuatro localidades tienen un clima semiárido (lluvias en torno a 250-300 mm), pero las localizadas a menores altitudes tienen un mayor déficit de presión de vapor debido a su mayor temperatura promedio anual López 2000. En Resultados se brinda una explicación más detallada de la flora y vegetación de estas cuatro localidades.
Los patrones espaciales se determinaron mediante un mapeo de las posiciones de los arbustos presentes en las parcelas de estudio. El índice conocido como función g fue el que elegí para determinar los patrones espaciales de los arbustos (ver López et al. 2010, 2017, para más detalles). De forma sencilla, mediante este análisis el software permite delimitar círculos de diámetro creciente en torno a cada uno de los arbustos mapeados. El borde del círculo presenta una especie de banda o anillo de grosor ajustado a la densidad de plantas de la parcela (esto suele establecerlo el software automáticamente). Se cuentan todos los puntos (arbustos) que estén presentes dentro de ese anillo. El programa repite este proceso para todos los puntos y para diferentes diámetros (escalas) del círculo. El mismo programa promedia estos recuentos de las diferentes escalas y esto nos da el valor observado de la función g de nuestra parcela. Si ese valor es igual a uno, entonces el patrón no es diferente de un patrón aleatorio. Si es mayor que uno, entonces es agregado, y si es menor que uno, uniforme. Esto es así para un mismo tipo de puntos (por ejemplo, todos los arbustos de Bulnesia). Cuando se trata de dos tipos de puntos (por ejemplo, arbustos de Bulnesia frente a arbustos de otra especie), si el valor de la función g es mayor que uno, entonces tenemos atracción entre los dos tipos de puntos; si el valor de g es menor que uno, hay repulsión o segregación de los puntos. Para tener certeza estadística de que el patrón es realmente diferente de uno se realizan múltiples simulaciones (llamadas simulaciones Montecarlo) en las cuales el programa reasigna coordenadas a los puntos (arbustos). Esta reasignación se realiza mediante diferentes tipos de restricciones a la manera en que se asignan las nuevas coordenadas a los puntos. Estas restricciones tienen detrás una lógica biológica. A esto es a lo que se conoce como modelo nulo. Según la diferente lógica biológica tendremos un modelo nulo diferente. Gracias a estos modelos nulos se generan unas bandas (bandas/envueltas de simulación o intervalos de confianza) cuyos límites superior e inferior quedan por encima/debajo de una recta trazada a partir del valor de g = 1 (el valor aleatorio). Cualquier valor que esté dentro de esta banda de simulación no difiere del valor de g = 1 de manera significativa, y por tanto se considera aleatorio. En consecuencia, los modelos nulos son los que permiten poner a prueba la potencial existencia de un proceso dado (para más detalles sobre modelos nulos, ver Wiegand & Moloney 2004,2014,López et al. 2016,2020.
Tabla 1. Composición y abundancia de especies en las cuatro comunidades estudiadas. a. Cruzhuasa 2 (parcela de 50 x 20 m); b. Cruzhuasa (parcela de 20 x 10 m); c. Culpina (parcela de 30 x 20 m); y d. Alto Impora (no se realizó mapeo de puntos en parcela). En los tres primeros casos, la densidad se estandarizó a 1000 m2, ya que las parcelas diferían en área. Para Alto Impora se muestran los arbustos encontrados a lo largo de un transecto lineal (no hubo referencia de área; por eso hablo de densidad relativa). La cobertura absoluta de la vegetación, determinada mediante línea de intercepción y reflejada en términos de porcentaje, fue de 20, 25, 20 y 45, respectivamente, para Cruzhuasa 2, Cruzhuasa, Culpina y Alto Impora.

La población de Cruzhuasa 2 es la mejor fuente de información para Bulnesia, pues en ella mapeamos las coordenadas de 438 individuos de todas las edades (plántulas hasta arbustos viejos). Allí estudiamos el patrón de todos los individuos (modelo nulo CSR, por sus siglas en inglés complete spatial randomness) o de aleatoriedad espacial completa, y con el cual se puede determinar si el patrón espacial es agregado, uniforme o aleatorio), el CSR de varias clases de tamaño, el modelo nulo de independencia (que permite evaluar asociaciones interespecíficas), el modelo nulo de condición antecedente (con el cual se puede evaluar la potencial existencia de facilitación) y el modelo nulo de etiquetado o marcaje aleatorio (random labelling), que permite evaluar los patrones de mortalidad y la existencia de selfthinning (a veces traducido como autoaclareo), que es la muerte que ocurre debido a competencia intraespecífica en poblaciones de especies de plantas abarrotadas de individuos Harper 1977, Westoby 1984, Kenkel 1988.
Brevemente explico los diferentes modelos nulos (ver Wiegand & Moloney 2004ode la Cruz Rot 2006 para una descripción detallada). En el modelo CSR, la simulación Montecarlo asigna aleatoriamente coordenadas a los puntos sin restricciones, salvo la de que dos puntos no pueden caer en el mismo lugar. En los modelos nulos bivariados hay mayor complejidad. En el modelo de independencia, la estructura de los dos tipos de puntos se mantiene (es decir, las distancias relativas entre los puntos), pero una de estas estructuras se mueve (se rota, lo que se conoce como desplazamiento toroidal) respecto a la otra, de modo que se rompa la dependencia espacial entre los dos patrones. Cada rotación representa una nueva asignación de coordenadas, y por tanto un nuevo patrón aleatorio. En el modelo de condición antecedente se mantiene fijo el patrón de uno de los dos puntos y las posiciones del segundo patrón de puntos se asignan al azar (mediante un CSR). Este modelo nulo pone a prueba la existencia de más puntos de una determinada clase (por ejemplo, plántulas) en las cercanías de los puntos de otra clase (por ejemplo, arbustos). En el modelo de etiquetado aleatorio, un primer proceso genera el patrón de puntos y un proceso posterior asigna marcas a los puntos. Esto significa que se mantienen fijas las posiciones de los dos tipos de puntos, y lo que se asigna son marcas o etiquetas a cada uno de ellos (por ejemplo, la etiqueta de muerto o vivo). Para un a = 0,05, normalmente se realizan 199 simulaciones, y se eliminan los valores de la función g de las cinco simulaciones con los valores más altos y de las cinco con los valores más bajos.
Otra manera de abordar el asunto del selfthinning, aunque más indirecta y menos precisa, es mediante un estudio de los patrones espaciales por clases tamaño. El tamaño es más indicado que la edad cuando se quiere establecer la estructura espacial de muchas especies, ya que está relacionado de mejor manera con sus tasas vitales Begon 1996. Se esperaría que el patrón pasara de agregado a aleatorio y finalmente a uniforme conforme aumenta el tamaño de los individuos. Este tipo de análisis se realizó en Cruzhuasa 2 y, en parte (el tamaño de muestra era inferior), en Cruzhuasa. Para ello se utilizaron la altura y el área de la copa como parámetros de tamaño, ya que no está claro cuál de estas variables representa mejor el estado de desarrollo de Bulnesia. Dada la forma de crecimiento de la especie (plantas muy bajas con copas muy extendidas), quizás este parámetro podría dar mejor idea de cuánto afecta el tamaño a los patrones espaciales. Para ellos utilicé la fórmula de la elipse (n * 1/2a * 1 /2b, donde a y b son los diámetros mayor y menor, respectivamente). Dado que solo Cruzhuasa 2 presentaba abundancia de individuos en todas las clases de tamaño, este análisis se llevó a cabo solo para esta localidad. La separación de los individuos en clases de tamaño a partir del tamaño de copa es menos intuitiva (áreas), de modo que arbitrariamente generé cuatro clases de diámetro: plántulas, saplings (brinzales), jóvenes y adultos. Los cien individuos más pequeños los asigné a la primera clase, los cien sucesivos a la siguiente, los noventa y nueve siguientes pasaron a la clase jóvenes, y los últimos noventa y nueve, a la clase de adultos). Este enfoque de separar los individuos en clases de tamaño permitió además hacer una estimación de la estructura poblacional de Bulnesia. Por otro lado, como el mapeo se realizó en parcelas (y por tanto había una referencia de área), se pudo estimar la densidad absoluta y relativa de Bulnesia en cada una de las localidades estudiadas (en Impora Alto se pudo determinar solo la densidad relativa). La densidad absoluta indica el número de individuos de Bulnesia en relación con el área de referencia (1.000 m2). La densidad relativa nos da el porcentaje de individuos de Bulnesia en relación con todos los individuos de la parcela (todas las especies). En las otras localidades se realizaron los mismos análisis, salvo aquellos para los que el tamaño de muestra era bajo. Los patrones espaciales se determinaron mediante el software llamado Programita Wiegand & Moloney 2004,2014.
Resultados
Se encontraron poblaciones de la especie solo en la región de los Cintis, Chuquisaca. Comienzo la descripción con Cruzhuasa 2, que es donde encontré la mayor cantidad de individuos de Bulnesia. Cruzhuasa 2 fue la la comunidad donde Bulnesia tuvo mayor abundancia, tanto en términos de cobertura relativa como de densidad relativa (casi el 50% en ambos casos). Además, aquí se encontraron los individuos de Bulnesia de mayor tamaño (Tabla 1a).
Tabla 2 Estructura poblacional de tres de las comunidades vegetales estudiadas. Las clases de altura van de 1-9 cm, de 10-19.9 cm, de 20-29.9 cm, y así, sucesivamente.
| Clase de tamaño (altura; cm) | Cruz Huasa | Cruz Huasa (%) | Número de muertos | Cruz Huasa2 | Cruz Huasa2 (%) | Número de muertos | Culpina | Culpina (%) | Número de muertos |
| < 10 | 4 | 0.03 | 1 (25%) | 37 | 0.08 | 3 (8%) | 1 | 0.01 | |
| 10-20 | 17 | 0.14 | 7 (41%) | 83 | 0.19 | 4 (5%) | 1 | 0.01 | 1 |
| 20-30 | 50 | 0.41 | 12 (24%) | 99 | 0.23 | 8 (8%) | 4 | 0.05 | |
| 30-40 | 28 | 0.23 | 5 (18) | 76 | 0.17 | 7 (9%) | 13 | 0.15 | 1 |
| 40-50 | 11 | 0.09 | 1 (9%) | 69 | 0.16 | 2 (3%) | 21 | 0.25 | 1 |
| 50-60 | 8 | 0.07 | 0 | 31 | 0.07 | 0 | 13 | 0.15 | 2 |
| > 60 | 4 | 0.03 | 1 (25%) | 43 | 0.10 | 0 | 31 | 0.37 | |
| TOTAL | 122 | 1.00 | 27 | 438 | 1.00 | 24 | 84 | 1.00 | 5 |
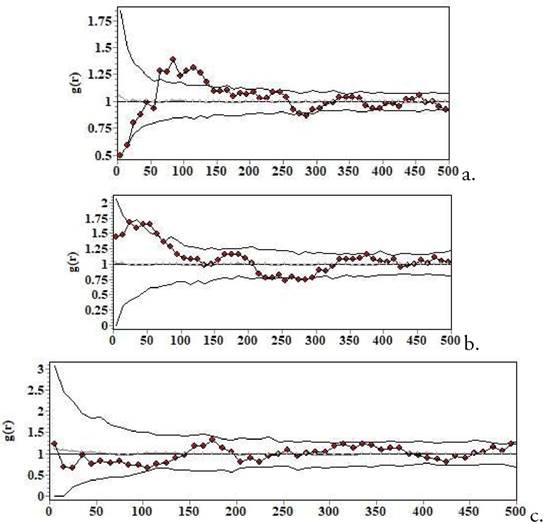
Figura 1. Patrones espaciales de todos los individuos de Bulnesia rivas-martinezii. a. Cruzhuasa 2; b. Cruzhuasa; c. Culpina. El eje de las x señala la distancia en cm. La línea punteada representa el patrón espacial observado. Las líneas grises representan la banda o envuelta de simulación (los intervalos de confianza). Cuando la curva punteada queda por encima de la línea gris, esto indica agregación de los individuos en las distancias en las que se produce ese rebasamiento, como en a. y b. Modelo nulo CSR o de aleatoriedad espacial completa.
Todas las clases de altura presentaron un buen número de individuos, aunque las clases intermedias fueron las que contaron con mayor densidad (Tabla 2a). Algo importante, encontré bastantes individuos muy jóvenes (plántulas/brinzales, es decir, seedlings/saplings). No obstante, esta clase de altura no fue la más numerosa. Aparentemente pocos individuos de Bulnesia mueren, y esto ocurre independientemente de la clase de tamaño (es decir, los porcentajes son bajos (menores que 10%) y distribuidos más o menos uniformemente entre las clases de altura).
El patrón espacial de Bulnesia en esta localidad fue agregado (entre 60 y 140 cm; Fig. 1a). Esto significa que en ese rango de distancias hubo más individuos en torno a un individuo cualquiera de la comunidad de lo que se esperaría por azar. Bulnesia presentó asociación positiva fuerte con Aristida sp. a escalas debajo de 50 cm. No se detectó interacción positiva ni negativa con Pseudosenegalia feddeana ni Parkinsonia andícola, dos de los mayores arbustos de la Prepuna (cuando se hizo esta comparación solo con los individuos de más de 30 cm de C. andicola, entonces se encontró repulsión a poco más de 2 m; comparar con el caso de Culpina, abajo). El patrón de mortalidad (análisis univariado de etiquetado aleatorio) de Bulnesia mostró que los grupos de individuos muertos estaban más agregados que el conjunto de los vivos y los muertos considerados como un todo (Fig. 2a). Este es un patrón esperado cuando existe selfthinning, y sugiere la existencia de algún tipo de competencia intraespecífica Kenkel 1988. No obstante, el resultado según el modelo nulo de etiquetado aleatorio bivariado no difirió de la aleatoriedad a ninguna de las escalas espaciales, lo que sugeriría que no hay competencia por recursos (Fig. 2b).
El análisis de los patrones espaciales por clases de alturas mostró agregación en las clases menores que 20 cm y en la comprendida entre 20 y 40 cm. En las clases más altas se percibió un patrón esencialmente aleatorio (no se muestran los resultados). En cambio, cuando empleé el tamaño de copa como indicador del tamaño de la planta, emergió un cambio gradual de patrón en función del tamaño: patrón agregado en las plántulas más pequeñas (plántulas), pasando por patrones cada vez menos agregados (brinzales y jóvenes) hasta patrones en los que hay repulsión a escalas debajo del metro (Fig. 3). Al igual que el patrón obtenido con el modelo nulo de marcaje aleatorio, esto sugiere algún tipo de competencia entre las plantas. Esto puede estar relacionado con la forma de crecimiento de Bulnesia, que parece favorecer el crecimiento horizontal de la copa que el vertical después de alcanzarse cierta altura.
En Cruzhuasa, la densidad de individuos de todas las especies en Cruz fue de casi 1.150 individuos, de los cuales 127 fueron de Br (11%). En términos de cobertura relativa, Bulnesia fue un poco más importante (15%). Esta es la comunidad que presentó individuos más pequeños de las cuatro estudiadas (Tabla 1b).
La estructura poblacional de Bulnesia en Cruzhuasa mostró predominancia de individuos en alturas intermedias, de manera bastante parecida al caso de Cruzhuasa 2 (Tabla 2b). A diferencia de aquella, se encontraron poquísimas plántulas. Los porcentajes de mortalidad fueron también bajos, e igualmente estuvieron algo más concentrados en alturas intermedias. Sin embargo, la escasez de plántulas sugeriría que la mortalidad es muy alta en este grupo de tamaño, o bien que el reclutamiento es muy limitado.
El patrón espacial de Bulnesia presentó débil agregación a escalas de entre 20 y 80 cm, es decir, a escalas un poco menores que en Cruzhuasa 2 (Fig. 1b). Bulnesia mostró dos picos de atracción con Flourensia fiebrigii a escalas debajo del metro (Fig. 4a). También hubo atracción con el cactus globoso Parodia maassii a escalas muy pequeñas (debajo de 50 cm, Fig. 4b) al emplearse el modelo nulo de independencia, pero no con el de condición antecedente (que es el que suele indicar facilitación). Se realizó un análisis de mortalidad, aunque con un tamaño de muestra muy inferior que el de Cruzhuasa 2 y que prácticamente no incluyó plántulas (que es donde mejor debería detectarse el patrón de mortalidad). Según este análisis, no hay evidencia de selfthinning en clases de tamaño por arriba del nivel juvenil en esta población. El patrón espacial de los individuos más viejos de la comunidad (iguales o mayores que 30 cm) mostró aleatoriedad a diferentes escalas salvo a escalas de 3.50-4.0 m, donde existió agregación que puede estar vinculada a la heterogeneidad de recursos (no se muestran los resultados).
En Culpina, la densidad y cobertura relativa de Bulnesia se acercaron a 10% en esta comunidad, es decir, aquí Bulnesia presentó sus valores de abundancia más bajos (Tabla 1c). Esta es la población aparentemente menos saludable, ya que la mayoría de sus individuos se encontraron en las clases de tamaño mayores (Tabla 2c). Prácticamente no encontré individuos jóvenes, lo que implica que los factores que impiden el reclutamiento en Bulnesia son particularmente marcados aquí.
La población de Culpina no mostró un patrón agregado ni uniforme a ninguna escala, a diferencia del caso de las poblaciones de las localidades de menor altitud (Fig. 1c). Dado que casi todos los individuos de Bulnesia en Culpina eran más altos que 30 cm, este análisis es comparable al que se hizo para las localidades anteriores para el caso de individuos viejos. Bulnesia se encontró ligeramente agregada a escalas de 20-30 cm con F. fiebrigii (Fig. 5a), y también con Pseudosenegalia feddeana (Fig. 5b), a diferencia de lo que ocurrió en Cruzhuasa 2 (donde no se vio ningún tipo de interacción). Por el contrario, Bulnesia mostró repulsión con Parkinsonia andicola a escalas debajo de 1 m (Fig. 5c), aunque hubo cierta agregación a escalas de 1.502 m.
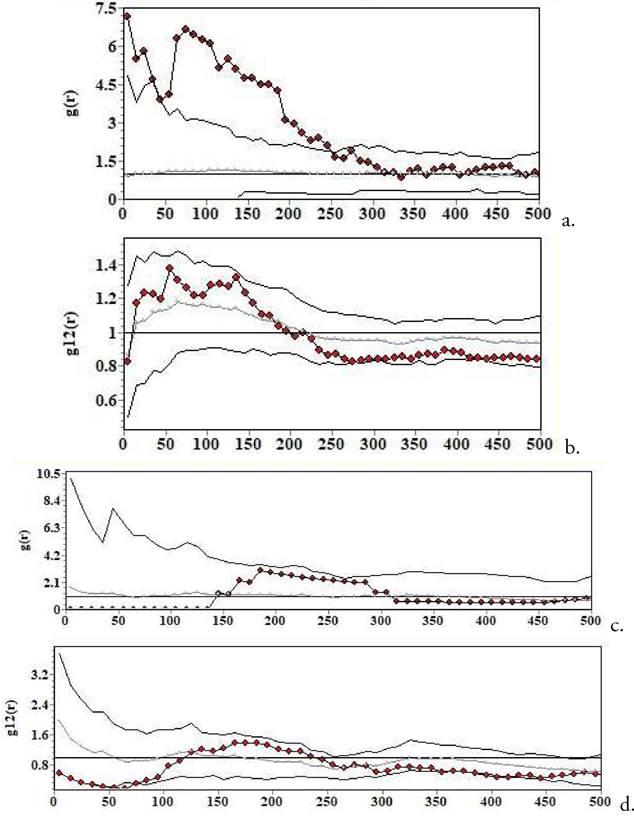
Figura 2. Patrones de mortalidad en Bulnesia rivas-martinezii en Cruzhuasa 2 (a. y b.) y Cruzhuasa c. y d.). El eje de las x señala la distancia en cm. La línea punteada representa el patrón espacial observado. Las líneas grises representan la banda o envuelta de simulación (los intervalos de confianza). a. y c. muestran el modelo nulo de etiquetado aleatorio univariado; b. y d. muestran el mismo modelo nulo, pero en su versión bivariada.
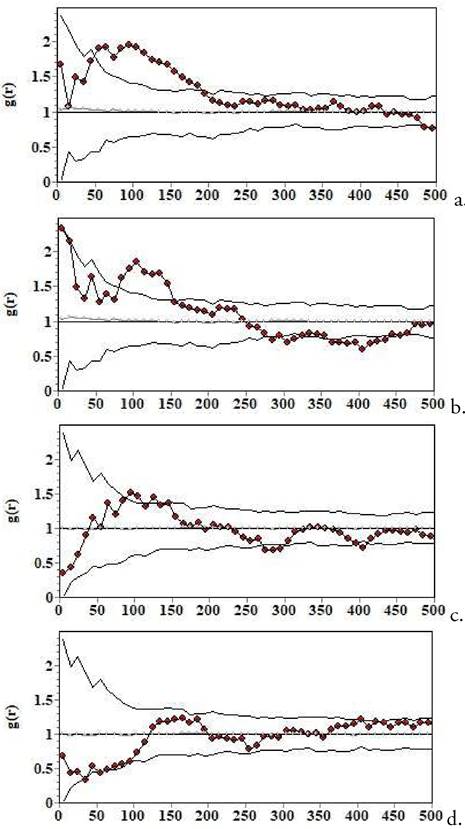
Figura 3. Patrón espacial de individuos basados en el área de sus copas: a. plántulas (seedling), b. brinzales (sapling), c. jóvenes, y d. adultos. El eje de las x señala la distancia en cm. La línea punteada representa el patrón espacial observado. Las líneas grises representan la banda o envuelta de simulación (los intervalos de confianza). Cuando la curva punteada pasa por encima de la línea gris, esto indica un patrón de agregación; cuando por debajo, un patrón uniforme. Nótense las diferencias con la Fig. 3 y el progresivo paso de patrones agregados a menos agregados, y finalmente a uniformes (en adultos), lo cual sugiere selfthinning. Modelo nulo CSR, o de aleatoriedad espacial completa.

Figura 4. Análisis bivariado de patrones espaciales que evalúa las interacciones entre Bulnesia rivas-martinezii y Flourensia fiebrigii a. y B. rivas-martinezii y el cactus globoso Parodia maassii, b. en Cruzhuasa. El eje de las x señala la distancia en cm. La línea punteada representa el patrón espacial observado. Las líneas grises representan la banda o envuelta de simulación (los intervalos de confianza). Cuando la línea punteada queda por encima de la gris, esto indica atracción entre las dos especies. Modelo nulo de independencia, o de desplazamiento toroidal.
En Alto Impora fue otra comunidad donde Bulnesia tiene gran importancia en cuanto a abundancia: 35% de la cobertura relativa y casi 30% en términos de densidad (Tabla 1d). Como en Cruzhuasa 2, Bulnesia fue la especie dominante. Lamentablemente, en esta localidad no se pudieron establecer parcelas para determinar patrones espaciales.
Discusión
Bulnesia rivas-martinezii tiene poblaciones relativamente grandes en los lugares donde está presente (valles de Cinti, Chuquisaca). Su cobertura relativa oscila entre 10 y 50% (Tabla 1). Sin embargo, su distribución parece ser parchosa (patchy; no está presente en todas las comunidades que visitamos dentro de su área de distribución). Si a esto añadimos el hecho de que el área de distribución de Bulnesia es muy restringida, su estatus de conservación debería ingresar en alguna categoría de amenaza.
Basado en los patrones espaciales de Bulnesia podemos concluir que la especie suele tender a la agregación (Fig. 1). Una de las causas podría ser la limitación a la dispersión. Esto se ve respaldado por la distribución aparentemente discontinua de la especie, y en general por su muy restringida área de distribución. Asimismo, la semilla no presenta adaptaciones aparentes a ningún tipo de agente dispersor (R.P. López, obs. pers., ver tambien Navarro 1994, en su Fig. 1), por lo cual una vez que se desprende del fruto queda debajo de la planta madre (el agua podría arrastrarla unos centímetros o pocos metros más allá del lugar de caída). Es interesante notar que los picos de agregación ocurren a una cierta distancia de la base de los arbustos. Esto podría deberse a la forma de crecimiento de Bulnesia: arbustos achaparrados y con la copa muy extendida, de modo que las flores/frutos se desarrollan a una distancia de la zona de enraizamiento de la planta, que sería la zona donde justamente caen las semillas. En Cruzhuasa la agregación va desde los 40 hasta los 60 cm, mientras que en Cruzhuasa 2 (donde los arbustos tienen copas mayores), esa distancia va de 60 a 130 cm.
La autoecología de Bulnesia no parecería ser la responsable de la distribución restringida de la especie, pues esta se presenta a diferentes altitudes dentro de los Cintis, lo que implica diferencias climáticas y, probablemente, edáficas de cierta importancia (más húmedad relativa, y posiblemente precipitación, más arriba). Si bien en Culpina no se encontró este patrón de agregación, hay que tener presente que esta población está compuesta por individuos mucho más viejos, en los cuales debería haberse producido ya selfthinning (ver siguiente parágrafo).

Figura 5 Análisis bivariado de patrones espaciales que evalúa las interacciones entre Bulnesia rivas-martinezii y Flourensia fiebrigii (a.), B. rivas-martinezii Pseudosenegalia feddeana (b.) y B. rivas-martinezii y Parkinsonia andicola (c.) en Culpina. El eje de las x señala la distancia en cm. La línea punteada representa el patrón espacial observado. Las líneas grises representan la banda o envuelta de simulación (los intervalos de confianza). Cuando la línea punteada queda por encima de la gris, esto indica atracción entre las dos especies; cuando queda por debajo de la línea gris, esto indica repulsión. Modelo nulo de independencia, o de desplazamiento toroidal.
Los patrones de mortalidad (ver Kenkel 1988: Wiegand & Moloney 2004,2014 para los detalles de este tipo de análisis) encontrados en Cruzhuasa 2 apuntan en principio a la existencia de selfthinning en esta población (Fig. 2). Empero, hay que considerar que el patrón de mortalidad abarcó individuos muertos de todos los tamaños, lo que sugiere competencia entre individuos de diferentes edades. Sin embargo, el análisis de los patrones espaciales por clases de tamaños de copas apoya las conclusiones del análisis de mortalidad con el modelo nulo de etiquetado aleatorio (caso univariado), es decir, sugiere también la existencia de selfthinning (Fig. 3). El análisis de etiquetado aleatorio bivariado, sin embargo, no muestra segregación de muertos y vivos, que es lo que se esperaría de la competencia por recursos (pero hay que considerar que el tamaño de muestra de los individuos muertos no fue muy grande). Los individuos posiblemente compiten por espacio, ya que la forma de crecimiento con la copa muy extendida hace que cada individuo requiera un área de superficie más importante que en otras especies. Otro indicador aún más indirecto de selfthinning es el de que los individuos más pequeños no son los más numerosos. Para tener mayor certeza de selfthinning en plántulas, necesitaríamos hacer este estudio solamente con individuos muertos de las clases de tamaño más pequeñas, y ello requeriría un tamaño de muestra grande de muertos en esta clase de tamaño/edad. Por ello, el número relativamente bajo de individuos en estas clases jóvenes también podría estar relacionado con otros procesos, como la baja producción de semillas viables, o baja germinación, o bajo establecimiento, o depredación (principalmente por cabras,) o una combinación de las anteriores. Por otro lado, el patrón de mortalidad de Cruzhuasa no indica selfthinning, pero el análisis en este caso estuvo basado en mucho menos individuos que en el caso de Cruzhuasa 2 y prácticamente no incluyó plántulas o juveniles, que es la etapa de la ontogenia donde más se debería detectar este fenómeno.
Bulnesia interactúa con algunas de las especies de los lugares donde habita (Figs. 4 y 5). El análisis de patrones espaciales muestra asociación positiva con una especie de cactus globoso, Parodia maassii (especie endémica de la Prepuna boliviano-jujeña), pero solo con el modelo nulo de independencia. Cuando se usa el modelo nulo de condición antecedente (específicamente diseñado para evaluar facilitación, Wiegand & Moloney 2014), la interacción desaparece, lo que podría indicar simplemente preferencias de hábitat similares en las dos especies. Otras especies leñosas de la Prepuna sí parecen actuar como potenciales nodrizas: Prosopis ferox y Cercidium andícola (López et al. 2007,Zenteno & López 2010). Esto no es extraño, dado que las plántulas de los cactus suelen ser muy sensibles a las altas temperaturas de los desiertos en que habitan Godínez- Álvarez et al. 2003,López et al. 2007. Zenteno & López (2010) encontraron que los arbustos que parecen ser más ventajosos para P. maassii son aquellos que no producen demasiada sombra, como ser los individuos de tamaños intermedios de P. ferox, los cuales tienen tamaños similares a los de adultos de Bulnesia, por lo cual sería interesante investigar más sobre las causas de esta asociación positiva.
Navarro (1994) caracteriza la distribución de Bulnesia como una que ocurre en zonas donde Pseudosenegalia feddeana es dominante, lo que indicaría una asociación atribuible a procesos biogeográficos. Resultó interesante verificar si hay alguna asociación atribuible a procesos ecológicos entre estas especies. Detecté una asociación positiva en Culpina pero no en Cruzhuasa 2 (en Cruzhuasa solo había dos individuos de P. feddeana). En principio podríamos concluir que estas dos especies también tienden a estar asociadas a escalas de individuo (o sea, la escala de las interacciones, Fig. 5b), ya que a escalas debajo del metro hay atracción. No obstante, a escalas un poco mayores (en torno a metro y medio, y por tanto todavía a escalas de individuo) hay más bien tendencia a repulsión. La atracción a escalas espaciales muy pequeñas podría tener que ver con facilitación de P. feddeana (árbol) hacia Bulnesia (arbusto), pero cuando realicé el análisis entre P. feddeana y plántulas de Bulnesia encontré más bien una tendencia a la repulsión. Entonces la asociación positiva podría tener que ver con preferencias compartidas de microhábitat, aunque estas se esperarían todavía a la distancia en la cual se encontró repulsión (el metro y medio).
También es difícil de explicar la relación positiva con una gramínea perenne, Aristida sp., o con otro arbusto característico y dominante de la Prepuna, Flourensia fiebrigii. En las dos comunidades donde Bulnesia y F. fiebrigii coocurren se ve esta asociación positiva, pero se necesitan más datos para poder especular sobre las causas de esta asociación. Es interesante la observación de que tanto Bulnesia como F. fiebrigii (muy resinosa) y Aristida sp. no parecen ser ramoneadas por el ganado caprino y vacuno (S. Beck, com. pers. 2023), lo cual podría sugerir un posible mecanismo que explique la asociación positiva. Sin embargo, por el momento es difícil especular más al respecto. Podríamos también conjeturar preferencias parecidas de microhábitat y ausencia de fuerte competencia por recursos. Por el contrario, Bulnesia suele mostrar repulsión con Parkinsonia andicola (Fig. 5c), pero cierta atracción a escalas intermedias (1.50-2 m), lo cual sugiere que estas dos especies compiten si están muy cerca, o bien hay una interferencia en el crecimiento de las copas (dada la forma de crecimiento de Bulnesia); sin embargo, la atracción podría sugerir algún beneficio para Bulnesia si esta crece cerca de P. andicola (por ejemplo, mayores niveles de nitrógeno en el suelo, López et al. 2007). Empero, Bulnesia y los individuos mayores que 30 cm de P. andicola (equivalentes a los individuos de P. andicola en Culpina) muestran repulsión en Cruzhuasa 2, lo que en principio muestra que las interacciones positivas pueden no ser una norma, y esto indicaría que son hasta cierto punto débiles. Cuando sí se producen, podrían estar asociadas a mejoras en el suelo, como ya se ha sugerido para otros desiertos (López et al. 2016). Esto podría ser así, visto que P. andicola es una de las mayores plantas de la Prepuna, y potencialmente una fuerte competidora, pero también P. feddeana es una especie con individuos arborescentes y, como se vio, tiende a tener una asociación positiva con Bulnesia a distancias muy cercanas a los arbustos.
Los estudios de los patrones espaciales ayudan a detectar la existencia de potenciales interacciones, pero al tratarse de enfoques de tipo observacional, no permiten que se puedan establecer relaciones causa-efecto definitivas Underwood 1997,Schob et al. 2013. Para ello es necesario el complemento de un enfoque más experimental, ojalá acompañado de ulteriores estudios observacionales derivados de inferencias que se puedan extraer de los patrones hasta ahora encontrados. Es importante que las biólogas y los biólogos bolivianos aceleremos el conocimiento de la ecología de nuestras especies y comunidades biológicas a partir de estas diferentes aproximaciones científicas, a fin de poder aportar con mayor conocimiento empírico (más allá del limitado pero muy empleado enfoque prueba y error) a la conservación, restauración y gestión de éstas.
Conclusiones
Se caracterizaron los patrones espaciales uni y bivariantes del endemismo boliviano Bulnesia rivas-martinezii (Bulnesia), así como su abundancia y estructura poblacional en su zona de distribución principal: la Prepuna boliviana. Bulnesia tiene una distribución discontinua, pero es comparativamente abundante donde está presente. La estructura poblacional difiere de localidad a localidad, aunque en todas ellas las clases intermedias tienden a ser más abundantes. Bulnesia tiende a presentar patrones agregados (más evidentes donde hay plantas jóvenes). Además, hay indicios importantes de la existencia de selfthinning. Bulnesia presenta interacciones positivas con varias especies de diferente hábito.

