











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
Las interacciones planta-animal desempeñan un papel fundamental dentro de los ecosistemas (Banerjee et al., 2022), estas interacciones determinan la coexistencia de las especies y la persistencia y estabilidad de las comunidades (CaraDonna et al., 2021).
En la vasta diversidad de interacciones que existen una de las más estudiadas es la interacción de planta-polinizador (Mitchell et al., 2009; Sargent & Ackerly, 2008; van der Kooi et al., 2021). La polinización zoófila es una interacción de tipo mutualista donde ocurre la transferencia de polen de las anteras de una planta al estigma de otra (Khalifa et al., 2021). Esta transferencia habitualmente depende de las abejas y otros animales como aves, murciélagos, escarabajos y polillas (Khalifa et al., 2021). Las plantas utilizan rasgos florales como tamaño floral (Lozada- Gobilard et al., 2023), color (van der Kooi et al., 2019), aroma (Bisrat & Jung, 2022), etc., como señales para atraer a los polinizadores y a cambio los polinizadores reciben una recompensa, normalmente alimento (néctar o polen) (van der Kooi et al., 2023; Minnaar et al., 2019; Zariman et al., 2022).
Las plantas con flores tienen diferentes estrategias para evitar su autopolinización diferenciando el desarrollo y posición de sus órganos sexuales (Shivanna, 2015). Por ejemplo, algunas poblaciones de plantas están compuestas por dos o tres morfotipos que difieren en las alturas a las que se sitúan los estigmas y las anteras en las flores (Yuan et al., 2023), esto se denomina heterostilia. Este polimorfismo se asocia frecuentemente a un sistema de auto incompatibilidad e incompatibilidad entre morfotipos (Barrett, 2019).
Al hablar de polinización, también se debe tomar en cuenta la transferencia de polen entre flores de diferentes especies (Streher et al., 2020). En la naturaleza, el estigma de una flor abierta está expuesto al polen de su propia especie y de especies cercanas y lejanas (Huang et al., 2023). Al traslado de polen de una especie A (flor donadora) al estigma de una flor de especie B (flor receptora), se denomina transferencia de polen interespecífica, esto genera competencia de las flores por los polinizadores (Morales & Traveset, 2008).
La transferencia de polen interespecífica tiene efectos negativos en la flor receptora de polen disminuyendo su adecuación (Arceo-Gómez et al. 2019). Además, existe una pérdida de polen de parte de la flor donadora, ya que el polen recogido por el polinizador se va quedando en flores de otras especies (Ashman et al., 2020; Morales & Traveset, 2008).
Comprender cómo se desplaza el polen es fundamental para entender la especiación, diversificación y evolución de las plantas con flores (Moreira-Hernández & Muchhala, 2019).
Por lo que, nos planteamos las siguientes preguntas: ¿La transferencia de polen está restringida por la especie? y dentro de una misma especie ¿la transferencia de polen está facilitada por la heterostilia? Si la transferencia de polen está restringida por la especie esperamos observar mayor transferencia de polen intraespecífica que interespecífica.
Y si la transferencia de polen está facilitada por la heterostilia esperamos observar transferencia de polen de flores brevistilias a longistilias y viceversa.
Materiales y métodos
Área de estudio y selección de especies
El estudio se realizó en uno de los caminos adyacentes a la Reserva Privada “Potrerillos del Güendá”, en Santa Cruz, Bolivia.
Se ubica dentro de una zona biogeográfica de transición entre las ecorregiones amazónica y chiquitana (Navarro et al. 1996).
Se definió un transecto de 500 m dentro del camino (17°40'37.18"S 63°26'1.01"O y 17°40'38.48"S 63°25'45.41"O) (Fig. 1A) y en base a la vegetación presente se seleccionaron dos especies de plantas con flores morfológicamente similares en cuanto a color y tamaño:
Pavonia sidifolia (Malvaceae) (Fig. 1B) y Turnera krapovickasii (Passifloraceae), esta última a su vez presenta heterostilia, con los dos morfotipos característicos, flores brevistilas: anteras por encima del estigma (Fig. 1C) y flores longistila: anteras por debajo del estigma (Fig. 1D).
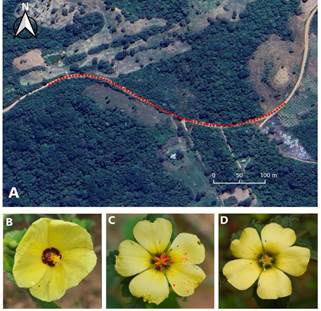
Fuente: (Fotos: (A) Mariana Zelada), (B) Sissi Lozada Gobilard, (C y D) Daniela Ramos)
Figura 1 (A) Transecto de estudio, camino adyacente a la reserva Potrerillos del Güendá. Especies seleccionadas para el estudio (B) P. sidifolia, (C) T. krapovickasii brevistila., (D) T. krapovickasii longistila. En las fotografías (C) y (D) pueden observarse análogos de polen
Movimiento de análogos de polen
Se definieron como parche a todos los brotes cercanos entre sí y se les asignó un código y número. Se utilizaron polvos fluorescentes como análogos del polen para observar si existía movimiento entre las dos especies de flores y entre morfotipos (Kearns e Inouye 1993).
Cada individuo recibió un polvo fluorescente de un color determinado: Amarillo para P. sidifolia (P), Naranja para T. krapovickasii brevistila (TB) y Verde para T. krapovickasii longistila (TL). Se colocaron polvos fluorescentes sobre la antera, utilizando un pincel de cerdas finas en aproximadamente 1/3 de las flores abiertas de cada parche.
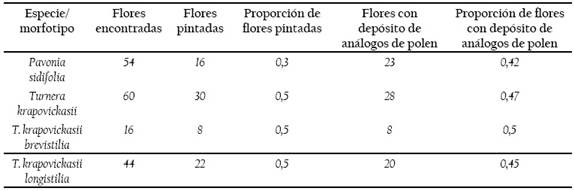
Al cabo de una hora se recogieron el total de las flores abiertas presentes en el transecto, pintadas y no pintadas (n= 54 flores de P. sidifolia, 16 de T. krapovickasii brevistila y 44 de T. krapovickasii longistila; Tabla 1). Las flores se inspeccionaron con un estereoscopio (AmScope SE306) y luz de linternas UV para una mejor visualización de los polvos fluorescentes.
De este modo, determinamos la proporción de flores que recibieron análogos de polen del mismo o de diferente color. Consideramos la presencia de análogos del polen en una flor como indicador de transferencia.
Análisis estadístico
Se elaboraron tablas de contingencia donde se incluyeron los pares de donadores y receptores. Una tabla consideró los pares de donador-receptor entre especies: T-P, T-T, P-T y P-P. Otra tabla consideró los pares de donador-receptor entre morfotipos de la misma especie con heterostilia: TL-TL, TL-TB, TB-TB y TB-TL.
Se realizó un modelo generalizado lineal nulo y un modelo generalizado lineal donde se incluyeron las interacciones entre los donadores y receptores de polen.
Mediante una prueba de Chi cuadrado se compararon los modelos generados. Los análisis estadísticos se hicieron en el ambiente R v. 4.2.2 (R Core Team, 2022).
Resultados
Se encontraron 54 flores de P. sidifolia y 60 flores de T. krapovickasii, de las cuales se encontraron 23 y 28 flores donde se depositaron análogos de polen respectivamente. De las 60 flores de T. krapovickasii, 16 eran flores brevistilas y 44 flores longistilas, de las cuales se encontraron 8 y 20 flores respectivamente con depósito de análogos de polen después de una hora de exposición. (Tabla 1).
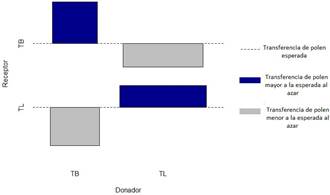
La transferencia de polen fue mediada significativamente por el factor especie (P= 7.786e- 08, DF= 1, X2= 28.859; Fig. 2). Se observó mayor transferencia de polen de lo esperado entre flores de pares donador-receptor de la misma especie (T-T, P-P). Por el contrario, se observó menor transferencia de polen de lo esperado entre los pares donador-receptor de diferentes especies (T-P, P-T). La transferencia de polen fue mediada significativamente por la heterostilia presente en flores de T. krapovickasii (P= 0.00194, DF= 1, X2= 9.6057; Fig. 3).
Sin embargo, hubo mayor transferencia de polen de lo esperado entre flores de pares donador- receptor con el mismo morfotipo (TL-TL, TB-TB). Y una menor transferencia de polen de lo esperado entre los pares donador-receptor con diferente morfotipo (TL-TB, TB-TL).
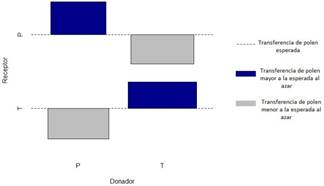
Figura 2 Transferencia de polen observada entre flores de dos especies T= T. krapovickasii y P= P. sidifolia, el ancho de las barras representa la proporción de flores de cada especie.
Discusión
El número de flores encontradas de T. krapovickasii y P. sidifolia fueron similares, por lo que el efecto de la abundancia no afectó a los resultados, posiblemente fueron otros los factores que afectan la transferencia interespecífica de análogos de polen.
Sin embargo, la diferencia entre la abundancia de flores longistilas y brevistilas pudo tener efecto en la transferencia de polen en las flores de T. krapovickasii. Estudios sugieren la tendencia de que a mayor abundancia de flores disminuye la transferencia de polen (Cohen et al., 2021; Monasterolo et al, 2022). Aún no se ha estudiado sobre cómo la diferencia en la abundancia de flores brevistilas y longistilas afecta la transferencia de polen.
La transferencia de polen interespecífica entre flores de T. krapovickasii y P. sidifolia fue menor de lo esperado. De manera similar, se observó menor transferencia de lo esperado de TL-TB y de TB- TL. Morales y Traveset (2008) reportaron la tendencia de los polinizadores de visitar las flores de un mismo individuo antes de pasar al siguiente individuo. Debido al tiempo de exposición en el estudio, es probable que los resultados reflejan esta tendencia, los polinizadores transfirieron el polen dentro del mismo parche o incluso dentro de la misma planta y no se movieron a otros parches.
Un factor que pudo haber afectado la transferencia de polen interespecífico es la distancia y la separación entre los parches de T. krapovickasii y P. sidifolia, los parches de T. krapovickasii se encontraban al lado izquierdo del transecto, mientras que los parches de P. sidifolia, en su mayoría, se encontraban al lado derecho. Estudios anteriores indican que la escala espacial y el grado de mezcla de flores de diferentes especies influyen en la transferencia de polen interespecífico (Morales & Traveset, 2008), no necesariamente tiene que ser una distancia muy grande, bastan metros para evitar la transferencia de polen interespecífica (Underwood et al., 2020).
El tiempo de espera de transferencia de polen durante este estudio fue de una hora. A pesar de que estudios como el de Brown & Mitchell (2001) indican que no es necesario tiempos muy largos para observar transferencia de polen interespecífico, es probable que el tiempo de espera haya sido corto para observar transferencia de polen interespecífica. Además, la transferencia de polen también pudo haber estado afectada por el clima, durante la hora de espera hubo intervalos de lluvia que influyeron en la actividad de los insectos.
Diferentes estudios observaron efectos negativos en la adecuación de las plantas cuando el polen interespecífico se depositó antes del polen conespecífico (Caruso & Alfaro, 2000) aunque esto depende de cada especie (Hao et al., 2023; Streher et al., 2020). Es probable que la transferencia de polen interespecífico durante esta primera hora reduzca las posibilidades de la existencia de estos efectos negativos en la adecuación de nuestras especies.
Además, está reportado que más que la deposición de polen heteroespecífico, el factor que tiene más impacto en las poblaciones de plantas es la pérdida de polen conespecífico (Morales & Traveset, 2008). Para entender cuál de estos dos factores afecta más a las poblaciones de estas dos especies es importante realizar estudios dirigidos a este tema.
En base a nuestros resultados, sugerimos realizar estudios donde se evalúe si existe transferencia intraespecífica de polen entre individuos o si la transferencia de polen está restringida entre flores de un mismo individuo, para esto proponemos marcar solo un individuo de cada especie y morfotipo, además el marcar a un solo individuo permitiría evaluar la distancia de transferencia.
En paralelo, sugerimos realizar estudios para identificar a los polinizadores de estas especies y evaluar qué polinizadores comparten estas dos especies morfológicamente similares.
La transferencia de polen estuvo restringida por la especie, pero no fue facilitada por la presencia de heterostilia, se recomienda evaluar la transferencia de polen incrementando el tiempo de exposición de los análogos de polen, evaluar el efecto de la distancia entre los parches de T. krapovickasii y P. sidifolia y además investigar más sobre los polinizadores que comparten para comprender más sobre la transferencia de polen intra e interespecífica en esta zona.

