











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
La relación entre las plantas con flores y los polinizadores rara vez ocurre uno a uno, dado que la mayoría de las especies de plantas son visitadas por más de una especie polinizadora (Ashworth et al 2015).
Por lo tanto, un aspecto central del estudio ecológico y evolutivo de las interacciones entre plantas - polinizadores es la identificación de los visitantes florales que ejercen el mayor efecto positivo sobre el éxito reproductivo de las plantas (Mayer et al 2011). Se espera que los visitantes que tengan el mayor impacto positivo sobre el movimiento de polen y la formación de frutos y semillas, actuarán como agentes de selección y moldearán la evolución fenotípica floral (Stebbins 1970).
Aunque la importancia de un polinizador depende de su efecto sobre la adecuación final de las plantas (Fenster et al 2004), rutinariamente en los estudios sobre las interacciones planta- polinizador, se utilizan medidas aproximadas de dicha adecuación final, tales como la frecuencia de visitas, el contacto con las piezas fértiles, la cantidad de polen removido, o la formación de frutos y semillas para inferir las consecuencias ecológicas y evolutivas de dicha interacción (e.g. Vázquez et al 2005; Tadey 2011; Ferreiro et al 2017; revisado en: Ne’eman et al 2010; Schupp et al 2017). Aunque estas medidas proporcionan una aproximación útil, de ninguna manera es una evaluación definitiva de la eficiencia del polinizador y de su efecto sobre el éxito reproductivo de las plantas (Waser et al 1996, Schupp et al 2017).
Recientemente, se han presentado propuestas teóricas y metodológicas que argumentan a favor de una estimación más precisa de la eficiencia de los polinizadores que incluyen vincular aspectos cuantitativos (cantidad de visitantes) y cualitativos (calidad de la visita) al momento de estudiar la interacción entre las flores y sus visitantes (Ne’eman et al 2010, Freitas 2013, Schupp et al 2017, Minnaar et al 2019, Valverde et al 2019, Baranzelli et al 2020).
Sin embargo, la evaluación del efecto sobre el éxito reproductivo de las plantas más allá de la frecuencia de interacción, del movimiento de polen y la formación de frutos y semillas, es decir sobre la calidad de la progenie, sigue siendo un desafío importante en la ecología de la polinización y está pendiente de una mayor investigación (Herrera 2000).
Incluir estos aspectos postcigóticos tales como los niveles y tiempos de germinación de semillas que provienen de diferentes visitantes resulta fundamental para entender la ecología y la evolución de las interacciones planta-polinizador.
La eficiencia en la polinización sobre la calidad de la progenie puede evaluarse, por ejemplo, sobre los niveles de formación de frutos y semillas (Baranzelli et al 2020), incluso es posible evaluar la diversidad genética de la progenie (Valverde et al 2019). Un aspecto menos valorado son los niveles de germinación de las semillas promovidos por los distintos polinizadores (Herrera 2000, Ne’eman et al 2010; Schupp et al 2017, Valverde et al 2019).
Esto podría ser un estimador clave en el desempeño de la polinización, porque brinda información de la contribución de diferentes polinizadores a la composición de individuos de la siguiente generación y del éxito reproductivo a largo plazo (Herrera 2000).
Si la eficiencia de un determinado visitante tiene consecuencias sobre los niveles de germinación de las semillas, sería posible determinar la importancia de aspectos cualitativos durante la polinización, tales como la calidad del polen, que reducen la depresión por endogamia e incremento de fenotipos favorables en la progenie (Valverde et al 2019).
Por ello, a pesar de ser un aspecto menos valorado, al momento de evaluar la eficiencia en la polinización, los niveles de germinación de semillas promovidos por diferentes visitantes florales, podrían darnos pistas claves sobre sus roles relativos sobre la evolución del fenotipo floral de las plantas (Stebbins 1970, Herrera 2000, Ne’eman et al 2010).
Lepechinia fioribunda (Lamiaceae) es un arbusto cuyas flores son polinizadas principalmente por abejas que toman néctar y colectan polen (Baranzelli et al 2020; Fig. 1). Observaciones y análisis del componente cualitativo y cuantitativo en la eficiencia de la polinización, han mostrado que dos de ellas, los abejorros nativos del género Bombus (B. opifex y B. pauloensis, de aquí en adelante Bombus spp) y las abejas melliferas exóticas Apis mellifera, resultan los polinizadores más eficientes (Baranzelli et al 2020; Fig. 1).
A pesar de su mayor eficiencia sobre el resto del ensamble de visitantes, entre estos dos grupos principales de polinizadores, también se observaron diferencias: tasas y duración de visitas a las flores fue más altas en A. mellifera, y mayor remoción y depósito de polen, y mayores niveles de formación de frutos y semillas en Bombus (Baranzelli et al 2020).
Al combinar los aspectos cuantitativos y cualitativos, se observó que los abejorros del género Bombus, resultaron los más eficientes y los que más contribuyen al éxito reproductivo de las plantas. Sin embargo, aún se desconoce cómo el comportamiento diferencial de estos dos grupos de polinizadores repercute en la calidad de la progenie, es decir, sobre los niveles de germinación de las semillas.
A la luz de las diferencias comportamentales en las flores de estos dos grupos de polinizadores, donde las abejas melíferas pasan más tiempo en las pero con un menor movimiento de polen, los abejorros realizan visitas más cortas pero más eficientes en cuanto al movimiento de polen, y las diferencias en la historia evolutiva de cada grupo de visitantes con la planta (Baranzelli et al 2020), el objetivo del presente estudio es evaluar una hipótesis específica que surge de dichas observaciones: las flores de L. fioribunda expuestas a regímenes de polinización contrastantes (esto es, abejorros nativos (Bombus spp) versus abejas melíferas exóticas (A. mellifera) deberían diferir en su contribución al reclutamiento de la población en la siguiente generación vía la calidad de la progenie.

Materiales y Métodos
Sistema y sitio de estudio
Lepechinia fioribunda (Lamiaceae) es un arbusto sub-perenne nativo de los bosques montanos en Argentina, Bolivia y Perú, comúnmente encontrado en hábitats abiertos y secos desde los 500 hasta los 3500 metros sobre el nivel del mar (Epling 1938).
Es una especie dominante que florece desde principios de octubre hasta finales de febrero, produciendo varias flores blancas hermafroditas bilabiadas por planta a lo largo de la temporada de floración (Fig. 1a-b).
Las flores están receptivas durante 1 día, presentan estigmas bífidos y cuatro óvulos. Son protándricas incompletas (las anteras maduran antes que los estigmas) y pueden autofertilizarse de manera autónoma (Roldan y Ashworth 2018). Los frutos maduran aproximadamente 3 semanas después de la polinización (Camina et al 2018).
El estudio se realizó en una población natural en la Reserva Natural Los Manantiales (31°9'40.34'' S, 64°21'03.67'' O) en el centro de Argentina, donde se estudiaron 33 individuos marcados durante la temporada de floración y fructificación 2018-2019.
Muestreo
Se utilizaron 33 plantas focales, en cada planta se marcaron entre 3 a 17 flores que no habían recibido visitas (256 flores en total), que luego de ser visitadas por Apis mellifera o Bombus spp fueron embolsadas. Un mes después del experimento de polinización, se registró la formación o no de frutos (113 frutos en total), y se colectaron las semillas de cada uno (363 semillas en total).
Germinación
Las semillas de L. fioribunda se sometieron a escarificación química con ácido sulfúrico (H2SO4) concentrado durante 30 minutos, un lavado con agua corriente durante dos minutos (Ashworth et al 2017), una inmersión de cinco minutos en suspensión fungicida ANTRAK 50 P.H ® (Agroquímica Tridente, 2019), un lavado de cinco minutos con jabón líquido comercial (30mL en 1L de agua destilada) y tres lavados con agua destilada estéril durante un minuto en la campana de flujo laminar.
Para cada tratamiento (Bombus spp y A. mellifera) se colocaron hasta cuatro semillas provenientes del mismo fruto, en cajas de Petri estériles de 6 cm de diámetro, con una base de agar bacteriológico (75%) y sacarosa (25%).
Cada caja se etiquetó y selló con parafilm, posteriormente todas las cápsulas se colocaron en una cámara de germinación en condiciones controladas de luz y temperatura (rango de variación diario: 25/15°C; con 12 h de fotoperiodo) donde el mayor valor de temperatura coincidió con las horas luz (Ashworth et al 2017) y se mantuvieron por 30 días bajo estas condiciones, hasta observar el crecimiento de las primeras hojas.
Se registró la cantidad de semillas germinadas por fruto, por planta y por tratamiento por cada día durante 30 días. Se obtuvo el porcentaje de germinación promedio por tratamiento, por planta y fruto para comparaciones posteriores.
Análisis estadístico
Para evaluar la eficiencia de polinización sobre la calidad de la progenie entre los dos grupos de polinizadores, se usaron modelos lineales mixtos generalizados (GLMMs; Zuur et al 2009) implementados en R v. 4.3.2 (R Core Team 2024). El modelo incluyó el porcentaje de germinación observado como variable respuesta (cantidad de semillas germinadas en relación al valor máximo de semillas que podrían formarse por fruto i,e. 4), el grupo de polinizador (Bombus spp y A. mellifera) como efecto fijo, y las plantas como efecto aleatorio dentro del grupo de polinizador.
La significancia estadística de los efectos fijos se estimó utilizando estimaciones de máxima verosimilitud restringida y comparaciones del criterio de información de Akaike (AIC). Se utilizó una distribución normal [family = gaussian] con la función lmer() (paquete: lme4; Bates et al 2015).
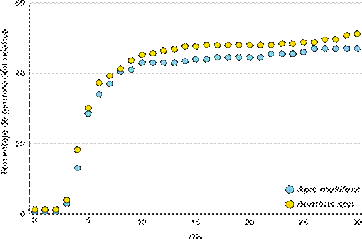
Para comparar si las curvas de germinación entre cada grupo de polinizador se diferenciaron a lo largo de los 30 días que duró el experimento, se evaluó si hubo diferencias entre las pendientes de regresión obtenidas del análisis de regresión lineal para cada grupo. Para obtener las pendientes de regresión se utilizó la función lm() en R, con el porcentaje de germinación acumulado como variable respuesta y el día como variable independiente. Para comprar las pendientes se utilizó la función diffslope() (paquete: simba; Jurasinski 2013) bajo 1000 permutaciones.
Resultados
El porcentaje de germinación medio de las semillas provenientes de flores polinizadas por Bombus spp fue en promedio (± SD) de 51.10% ± 40.43%, mientras que el porcentaje promedio de germinación de las semillas provenientes de flores polinizadas por A. mellifera fue (± SD) 47.00% ± 39.19 % (Fig. 2).
La comparación entre estos porcentajes de germinación indicó que no existía diferencias significativas (Estimate = 5.424; Standard Error = 7.057; t value = 0.769). Ambos grupos de polinizadores promovieron patrones acumulados de germinación similares en el tiempo (R2Bombus= 0.59; R2 A. mellifera = 0.60; Fig. 3). Concomitante, cuando se compararon las pendientes de germinación acumulada en función del tiempo, no se observaron diferencias entre ellas (Estimate = 0.02; Standard Error = 0.001; p value = 0.57)
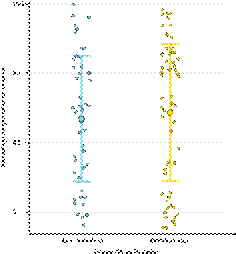
Figura 2 Porcentajes de germinación relativa por fruto observada en función de los dos grupos de polinizadores analizados para Lepechinia fioribunda (A. mellifera en celeste, y Bombus spp en amarillo). El punto de mayor tamaño representa el valor medio, las barras el desvío estándar y los puntos pequeños los frutos individuales. n = 113 frutos y 363 semillas.
Discusión
A lo largo de los años, se han utilizado diferentes indicadores de la eficiencia en la polinización de los distintos visitantes florales (Ne’eman et al 2010, Schupp et al 2017). En este trabajo, se evaluó la calidad de la progenie, vía tasas acumuladas y totales de germinación de semillas provenientes de flores polinizadas por uno de los dos grupos funcionalmente contrastantes de visitantes (abejas melíferas o abejorros) en L. fioribunda como indicador del éxito reproductivo de las plantas y el potencial aporte a la siguiente generación.
A pesar de las diferencias comportamentales, en abundancia, y las capacidades diferenciales de mover polen y formar semillas observadas previamente por parte de cada uno de estos grupos (Baranzelli et al 2020), no se registraron diferencias significativas en el potencial germinativo de las semillas. Estos resultados indicarían que la menor eficiencia de A. mellifera polinizando las flores de L. fioribunda (Baranzelli et al 2020), no afectaría, en términos relativos, el potencial de supervivencia de la progenie. A su vez, ilustran la importancia de cuantificar de manera completa, más allá de la polinización, la contribución de cada visitante floral al éxito reproductivo de las plantas (Schupp et al 2017, Valverde et al 2019).
A partir de las ideas sobre el principio del polinizador más eficiente de Stebbins (1970), se espera que los visitantes que tengan el mayor impacto positivo sobre el movimiento de polen y la formación de frutos y semillas, actúen como agentes de selección y moldearán la evolución fenotípica floral.
En el caso de L. fioribunda estudios previos indican que ese rol lo cumplen los abejorros nativos del género Bombus spp, dado que se observó un mejor ajuste fenológico entre la actividad de este grupo de visitantes y la maduración de las piezas fértiles de las flores y la producción de recompensas (Baranzelli et al 2020).
Asimismo, este mayor ajuste se refleja en la formación de frutos y semillas. Sin embargo, las abejas melíferas exóticas presentaron una sobre abundancia y a fuerza de ello, se observó que, al combinar aspectos cuantitativos y cualitativos, contribuyen significativamente al éxito reproductivo de las plantas (Baranzelli et al 2020).
Como corolario, se podría esperar que el banco de semillas en la población analizada esté compuesta principalmente por embriones provenientes de flores polinizadas por el visitante más eficiente, los abejorros nativos. Sin embargo, restaba la duda de si ese banco de semillas, diferencialmente compuesto, a su vez, podría ser afectado por mecanismos postcigóticos que promoverían, otra vez, la mayor eficiencia de los abejorros, el efecto selectivo de los rasgos florales, y la especialización del fenotipo floral a este grupo de polinizadores.
A pesar de ello, los resultados aquí observados muestran que los niveles de germinación del banco de semillas no se verían afectados por la calidad de las visitas de las abejas melíferas, o viceversa, tampoco se verían promovidos por la eficiencia de los abejorros. Estos resultados podrían explicarse por dos vías, la competencia gametofítica y la calidad de polen depositado por A. mellifera y Bombus spp.
Cuando el número de granos de polen depositado en un estigma supera el número de óvulos en el ovario puede existir competencia entre los micro gametofitos (teoría de competencia micro gametofítica; Mulcahy y Mulcahy. 1975, McKenna 1986, Davis et al 1987,). Tubos polínicos de crecimiento rápido llegan antes a fertilizar los óvulos que tubos de crecimiento lento, y los tubos de crecimiento rápido generan progenie más vigorosa. Esto se debe a que la mayoría de los genes del micro gametofito son heredados y expresados en el esporofito. Sin embargo, si la carga polínica en el estigma es baja, tubos polínicos de crecimiento rápido y lento llegaran a fecundar los óvulos.
Entonces, a pesar de que en L. fioribunda, los dos grupos de polinizadores analizados depositan diferentes cantidad de polen por visita, pero esa cantidad supera el número de óvulos (Baranzelli et al 2020), es probable que tanto Bombus spp. como A. mellifera generen niveles de competencia gametofítica similares, dando como consecuencia progenie de similar calidad.
La calidad de la progenie también puede ser afectada por el tipo de polen que le dio origen. Muchas especies de plantas presentan depresión por endogamia, esto es, el vigor de la progenie generada con polen de la misma planta (autopolinización), es menor que el vigor de la progenie generada con polen de otro individuo (polinización cruzada, (Angeloni et al 2011).
Estudios previos en L. fioribunda muestran efectos de depresión por endogamia en las defensas químicas contra herbívoros (Camina et al 2023). Entonces, podría ocurrir que L. fioribunda no muestre efectos de depresión por endogamia en la calidad de las semillas. .
De esta manera, en la última etapa del componente cualitativo en la eficiencia en la polinización, el del establecimiento de las plantas que componen la siguiente generación, la contribución a la calidad de la progenie sería similar entre estos dos grupos de polinizadores. Sin embargo, otros estudios han encontrado diferencias en las tasas de reclutamiento; por ejemplo, Herrera (2000) detectó que diferencias relativamente pequeñas en la composición y abundancia de grupos funcionales polinizadores tendría un impacto demográfico importante en las poblaciones de Lavandula latifolia (Lamiaceae).
Estos resultados, en combinación con los presentados en estudios previos para el sistema (Roldan y Ashworth 2018, Camina et al 2018, 2023, Baranzelli et al 2020), ilustran lo complejo que resultan las interacciones planta polinizadores, y el impacto y consecuencias diferenciales que pueden tener cada una de las etapas de la interacción (Schupp et al 2017).
Asimismo, resaltan la importancia de cuantificar de manera completa, más allá de la polinización, la contribución de cada visitante floral al éxito reproductivo de las plantas. Futuros estudios deberían evaluar el impacto a nivel genético en la composición de la progenie promovida por uno u otro grupo de polinizadores para tener una visión totalmente acabada de la eficiencia de los visitantes flores sobre el éxito reproductivo de las plantas (Valverde et al 2019).

