











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
INTRODUCCIÓN
Los camélidos sudamericanos domésticos (CSD): llama (Lama glama), alpaca (Vicugna pacos) (Stanley et al, 1994; Wheeler et al., 2006) y sus parientes silvestres la vicuña (Vicugna vicugna) y guanaco (Lama guanicoe), son recursos animales nativos estratégicos de la región Andina, todavía insuficientemente aprovechados desde la perspectiva de las oportunidades de mercado actuales y potencialidades hacia el futuro.
Su explotación constituye la mayor actividad para el sustento de familias en un vasto sector de la población Alto Andina, principalmente del Perú y Bolivia y en menor grado de Argentina, Chile y Ecuador (Fernández-Baca, 1991).
La crianza de llamas, específicamente, es una actividad concomitante a la de las alpacas, cuya implicancia económica es importante para un significativo porcentaje de la población alto andina, especialmente radicada en el Altiplano, debido a que produce fibra fina con características peculiares y carne de excelente calidad nutritiva, en comparación con otras carnes rojas.
Sin embargo, el sistema de producción actual utilizado por la mayoría de productores tiene muchas deficiencias de manejo tecnificado, lo que trae como consecuencia la disminución de la producción y productividad, debido a que las prácticas de crianza actuales son inadecuadas, por la que la calidad de las alpacas y llamas, sus productos están muy por debajo de su potencial (Wheeler, 2016). Uno de los múltiples factores que influyen en la producción en camélidos sudamericanos domésticos, y con mucho más énfasis en llamas, es el sistema de empadre que utiliza, el mismo que no es adecuadamente programado ni controlado y como consecuencia de ello trae un incremento de los niveles de consanguinidad en la población, generando una descendencia con mayor prevalencia de trastornos genéticos e inconvenientes sobre algunos caracteres productivos.
La consanguinidad (Inbreeding) en esencia, es el apareamiento entre animales que tienen uno o más antepasados en común (Wright, 1922; Cleveland et al., 2005; Carrillo y Siewerdt, 2010). Implica el apareamiento entre parientes cercanos, cuanto más cercano sea el parentesco entre ellos, mayor será la consanguinidad en la progenie resultante. Esta técnica reproductiva se suele utilizar para tratar de fijar o concentrar genes que controlan ciertos caracteres productivos en la progenie; sin embargo, la consanguinidad conduce a un incremento del nivel de homocigosis y como consecuencia aumenta la probabilidad de la aparición de efectos indeseables en el fenotipo, tanto en caracteres categóricos y cuantitativos, en lo último, como consecuencia de la depresión endogámica por consanguinidad, especialmente sobre la fertilidad, longevidad y en menor grado, en caracteres productivos. La medición de la consanguinidad se determina por el coeficiente de consanguinidad (F), que es la medida que hace referencia a la probabilidad que dos de los alelos de un individuo, ubicados en un mismo locus sean idénticos por descendencia (Falconer y Mackay, 1996).
No es pródiga la investigación científica de los efectos de la consanguinidad sobre diferentes caracteres en animales domésticos, la mayoría de ellos hace referencia al ganado bovino en diferentes razas (Fernández, 2005; Arzola, 2018; Burrow, 1998; Vergara, 2012; Verde, 2016). Estudios de los niveles de consanguinidad individual en poblaciones de camélidos sudamericanos domésticos, que permitan estimar la magnitud del efecto de la consanguinidad en los caracteres productivos son escasos, Vilela (2015) refiere un único estudio en alpacas, Mamani (2013) determina coeficientes de consanguinidad con propósitos descriptivos poblacionales únicamente; a la actualidad no se dispone de ningún referente de investigación con este propósito en variedades de la especie llama.
Sobre el particular, en el Centro de Investigación y Producción La Raya de la Facultad de Medicina Veterinaria y Zootecnia de la UNA-Puno, desde hace más de tres décadas, se ha implementado un módulo germoplasma de llamas para la valoración de sus cualidades productivas, los mismos que permitirán optimizar y planificar, con mayor racionalidad, las potencialidades bioproductivas de la crianza de esta especie animal en este espacio, en el que lamentablemente se evidencia ausencia clara de un aporte permanente de reproductores de fuera del hato.
Por esta razón, considerando la gran importancia de lograr mayor productividad sostenible en la crianza de llamas, se planteó el presente trabajo de investigación que estuvo dirigido a evaluar la magnitud de la consanguinidad individual del hato de llamas y su efecto sobre el peso al nacimiento y al destete, como caracteres que determinan el crecimiento en esta especie animal.
MATERIALES Y MÉTODOS
Ubicación de la zona de estudio
El Centro de Investigación y Producción La Raya se encuentra ubicado en el distrito de Santa Rosa, provincia de Melgar en el departamento de Puno; la ubicación geográfica expresada en unidades UTM son: 8 394 000.00 hasta 8 406 000.00 al Norte y 286 000.00 hasta 295 000.00 al Este. Geográficamente se encuentra entre los 13º00' y 17º18' de Latitud Sur y 71º08' y 65º50' de Longitud Oeste, con un patrón climático tipo "D", con temperaturas promedio que oscilan entre 9.5 y -4.2ºC, y una precipitación promedio anual 685.9 mm, a una altitud entre 4 100 a 5 000 m s.n.m. (Apaza, 2001); cuenta con una superficie de 5 095.87 ha.
Metodología
De los animales. Se utilizó datos generados por el hato de llamas del Centro de Investigación y Producción La Raya de la Facultad de Medicina Veterinaria y Zootecnia de la Universidad Nacional del Altiplano-Puno Perú, los que se recopilaron de los registros de parición, destete y empadre partir de año de 1984 hasta el 2019; luego de recopilados fueron compilados en una hoja electrónica del software EXCEL de Microsoft, por separado para ambas variedades de llamas (K'aras y Ch'akus). El número total de crías nacidas y consideradas para el estudio fue de 3 545, de los cuales 2 046 fueron llamas K'ara (1 012 hembras y 1 034 machos) y 1 499 llamas Ch'aku (726 hembras y 773 machos).
Procedimientos metodológicos. Los coeficientes de consanguinidad individual fueron determinados utilizando el procedimiento Proc inbreed del SAS (2001) (SAS/STAT 13.1) y software computarizado Pedigree viewer (Brian y Sandy Kinghorn, Universidad de New England, Australia), como procedimientos computarizados alternativos disponibles.
El efecto de la influencia del nivel de la consanguinidad sobre los caracteres peso al nacimiento y peso al destete se determinó utilizando en procedimiento de la regresión lineal simple del coeficiente de consanguinidad sobre el peso al nacimiento y peso al destete, separadamente, para ambas variedades de llamas utilizando el Software MINITAB 19. La significancia del factor coeficiente de consanguinidad sobre ambos caracteres, peso al nacimiento y peso al destete, fueron determinados utilizando un análisis de variancia, que para el caso de peso al nacimiento el modelo estadístico fue (Ecuación 1):
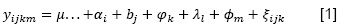
Donde: 𝑦𝑖𝑗𝑘𝑚 = peso vivo al nacimiento; 𝜇. . . = valor constante, igual a la media de la población; 𝛼𝑖 = efecto del año de producción; 𝑏𝑗 = efecto de año de la raza de la cría; 𝜑𝑘 = efecto del sexo de la cría; 𝜆𝑙 = efecto de la edad de la madre;𝑚 = efecto de Coeficiente de consanguinidad;𝜉𝑖𝑗𝑘 = error aleatorio.
Para el caso del peso al destete los datos originales fueron previamente ajustados a fecha única a través de la siguiente Ecuación 2:

Dónde: PEDEAJ = peso al destete ajustado a 210 días; PDESTT = peso al destete; PENAC = peso al nacimiento.
La significancia de la influencia del factor coeficiente de consanguinidad sobre el peso al destete fue determinado utilizando un análisis de variancia a través del siguiente modelo estadístico (Ecuación 3):

Donde:𝑖𝑗𝑘𝑚 = peso vivo al destete ajustad; 𝜇. . . = valor constante, igual a la media de la población; 𝛼𝑖 = efecto del año de producción; 𝑏𝑗 = efecto de año de la raza de la cría; 𝜑𝑘 = efecto del sexo de la cría; 𝜆𝑙 = efecto de la edad de la madre;𝜙𝑚 = efecto de coeficiente de consanguinidad; 𝜉𝑖𝑗𝑘 = error aleatorio.
RESULTADOS Y DISCUSIÓN
Coeficientes de consanguinidad
La Tabla 1 y Figura 1, muestran parte de valores de los coeficientes de consanguinidad individual para llamas de las variedades K'ara y Ch'aku, respectivamente, obtenidos por los procedimientos Inbreed del SAS y el procedimiento Pedigree Viewer, respectivamente.
Tabla 1 Coeficientes de consanguinidad individual en llamas de la variedad K'ara, por el procedimiento metodológico Inbreed del SAS.
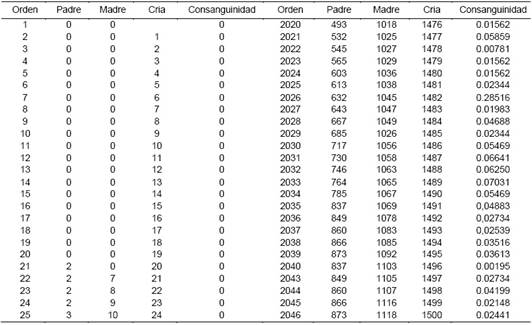
Figura 1 Coeficientes de consanguinidad individual en llamas de la raza Ch'aku, por el procedimiento metodológico Pedigree Viewer.
La consanguinidad desde una perspectiva estadística es definida como la probabilidad de que dos alelos en un mismo locus sean idénticos por descendencia, o sea, procedan de un ancestro común (Malecot, 1948); es el resultado del apareamiento entre individuos emparentados, es decir, que comparten uno o más ancestros en común; la consanguinidad, como efecto, tiene probada influencia negativa sobre la mayoría de características de importancia económica de los animales domésticos, denominado como depresión endogámica.
La Tabla 2 presenta valores de los coeficientes de consanguinidad para el número y porcentaje de llamas de las variedades K'ara y Ch'aku pertenecientes al Centro de Investigación y Producción La Raya de la Universidad Nacional del Altiplano-Puno, para cada valor de los coeficientes de consanguinidad determinados. Precisando que, el número de individuos evaluados en cada variedad es diferente, siendo mayor para animales de la variedad K'ara y menor para los de la variedad Ch'aku.
Tabla 2 Valores de coeficientes de consanguinidad según número y porcentaje de llamas, según variedad del Centro de Investigación y Producción La Raya UNA Puno.

Una apreciación notable que es evidente en la Tabla 2, es la existencia de un elevado número de individuos no consanguíneos en ambas variedades, el 40.13 y 34.00 % para las variedades K'ara y Ch'aku, respectivamente. El 55.77 % tienen un coeficiente de consanguinidad bajo, entre 0.0015 y 0.2500 en llamas de la variedad K'ara, un 58.04 % tienen este coeficiente bajo entre 0.0156 y 0.2500 en las de la variedad Ch'aku, respectivamente; sin embargo, es importante precisar que, dentro este último grupo, existe un número relativamente alto de individuos cuya consanguinidad individual es de 0.2500, en ambas variedades: 29.62 y 53.56 % para Kara y Ch'aku, respectivamente; el resto de valores corresponden a un relativo escaso número de animales, cuyos coeficientes de consanguinidad son 0.5 en ambas variedades, que representan el 4.10 y 7.96 % para K'ara y Ch'aku, también correspondientemente.
La mayor proporcionalidad de llamas con valores bajos y muy bajos, respecto del otro grupo con menor frecuencia y con valores del coeficiente de consanguinidad relativamente altos, se basa fundamentalmente en la poca profundidad del pedigrí, condicionado fundamentalmente por las características fisiológico-reproductivas de esta especie, especialmente hembras, que dejan poca descendencia en su vida productiva.
En este contexto, es necesario puntualizar que, en el proceso de determinación de los coeficientes de consanguinidad, mientras mayor información de generaciones pasadas se tiene, mayor es la probabilidad de encontrar coeficientes de consanguinidad mayores a cero; paradójicamente, en alguna medida no se satisface en el presente caso, dado que del total de llamas de la variedad K'ara (2 046), 595 llamas de ambos sexos fueron utilizados de manera repetida, generando 2 621 registros de individuos; de manera similar, cuotas diferentes de llamas machos y hembras fueron registrados reiteradamente en la determinación de del coeficiente de consanguinidad, generando un número de registros de 1 197 para machos y 1 424 para hembras en llamas K'ara; para el escenario de llamas de la variedad Ch'aku, con una población de 1 499 llamas de ambos sexos, 475 llamas de esta variedad fueron registradas reiteradamente en la determinación de coeficientes de consanguinidad, generando un total de 1 974 registros individuales, de los cuales 935 fueron machos y 1 039 fueron hembras (Tabla 3).
Estos valores indican que, a pesar de que se usó un número relativamente grande y diferente de reproductores de manera reiterada, especialmente machos, no generaron profundidad al pedigrí, que justifica el hecho de la ocurrencia de un gran número de llamas con un coeficiente de consanguinidad igual a 0.000. A este contexto es sumamente importante adicionarle el efecto de causas como la edad de la vida reproductiva útil de esta especie animal, la misma que no es muy prolongada y limita el uso de reproductores de ambos sexos, en numerosas generaciones de reproducción.
Los elevados porcentajes de llamas, 40.13 y 34.00 % para K'aras y Ch'akus, que registran coeficientes de consanguinidad individual igual a 0.000, para un período de estudio de más de tres décadas, fueron definidos por el escaso uso de reproductores en condiciones de antepasado común; un comportamiento similar fue reportado por Mamani- Cato et al. (2016), para un lapso de tiempo relativamente menor, indicando que el coeficiente de consanguinidad en llamas de 1993 al 2002 fue cero, debido a la compra frecuente de reproductores machos y hembras con la pertinente ausencia de registros genealógicos.
En la Tabla 2, además se aprecia un porcentaje considerable de llamas, el 16.52 % (K'ara) y 31.09 % (Ch'aku), tienen coeficientes de consanguinidad de 0.2500, como producto de apareamientos entre Abuelo-nieta o entre medios hermanos, que serían los tipos de apareamientos más frecuentemente ocurridos en este hato de llamas; un porcentaje menor de llamas (4.10 %) K’ara y un valor ligeramente superior (7.96 %) en llamas de la variedad Ch'aku, tiene un coeficiente de consanguinidad máximo de 0.50, esto como producto de un apareamiento más estrecho de padre con cría o madre con cría. Según Benjumea et al. (2010) pueden ser clasificados como endogamia estrecha (resultado del apareamiento de hermano con hermana, de padre con hija y de hijo con madre) y endogamia familiar (resultado de la unión entre medios hermanos, primos entre sí, tíos con sobrinos, etc.).
La Tabla 3, de manera similar, presenta dos parámetros importantes: a) la consanguinidad promedio poblacional para llamas de la variedad K'ara (0.010) y la correspondiente a la de las llamas de la variedad Ch'aku (0.005); los mismos que implican la probabilidad promedio, para la población de llamas, de que dos alelos de un mismo locus sean idénticos por ascendencia o a un antepasado común, valores que no implican ningún riesgo por la magnitud de los mismos, que está muy por debajo de 0.10 % valor considerado de cuidado en otras especies pecuarias. Mamani-Cato et al. (2016) para un similar hato de llamas refieren un valor de 0.11 % (0.0011) valor muy similar al encontrado en el presente estudio, dado las características poblaciones similares de llamas.
Valores referenciales similares fueron reportados en alpacas, cuyos valores fueron de 0.1654% (Vilela y Asparrin, 2013) y 0.04% (Mamani, 2013). b) La coascendencia promedio poblacional para llamas de la variedad K'ara (0.018) y para llamas de la Ch'aku (0.012), la que se define como la probabilidad de que los dos alelos de un mismo gen sean idénticos por descendencia, en otras palabras, es el porcentaje de pares de genes homocigóticos que el individuo posee por encima del promedio; aquí que el coeficiente de consanguinidad (F) es útil para medir el parentesco entre padres (coascendencia). La Tabla 3. presenta el número de individuos implicados para su determinación para cada variedad de llamas.

Los valores de los coeficientes de consanguinidad encontrados en el presente estudio, además de valores como el de 0.000 que contempla la ausencia de genes homólogos, así como de valores (0.0015, 0.039, 0.0078) insignificantes, estos como consecuencia de tipos de relaciones de apareamientos entre parientes no cercanos, no incluyen a los referidos por Arzola (2018) (Tabla 4). Sin embargo, es manifiesto la presencia valores del coeficiente de consanguinidad más elevados como consecuencia de tipos de relaciones de apareamientos, dada a la profundidad del pedigrí, especialmente en llamas de la variedad K’ara, y no así en llamas de la variedad Ch’aku cuyos valores de los coeficientes de consanguinidad todos están incluidos en la Tabla 4.
Tabla 4 Relación de apareamientos coeficiente y porcentajes de endogamia.

Adecuado de: Arzola (2018)
Existe escaso número de referentes científicos respecto de la determinación de coeficientes de consanguinidad individual y/o poblacional en la especie llama que permita contrastar resultados encontrados en la presente investigación. Al respecto Mamani-Cato et al. (2016) refieren un valor de 0.11 % para el coeficiente de consanguinidad promedio para llamas del Instituto Nacional de Innovación Agraria Perú, valor similar a los encontrados en el presente estudio, el que se encuentra dentro del rango determinado para llamas de las variedades K’ara y Ch’aku, con la restricción de que no fue frecuente ejecutar cruzamientos inter raciales, y consecuentemente no es viable ejecutar la determinación del coeficiente de consanguinidad que incluyan ambas variedades.
Existen estudios que determinan el coeficiente de consanguinidad en la especie alpaca, que pueden tomarse como referenciales para contraste con los determinados en el presente estudio; así, Mamani (2013) encuentra un valor promedio 0.04 % para el coeficiente de consanguinidad para la población de alpacas del banco de germoplasma de camélidos, Quimsachta (INIA-Puno), de manera similar, Vilela (2015) reporta un valor de 0.1654 % para el coeficiente de consanguinidad en población de alpacas del fundo Mallkini de la Empresa Michell, en ambos casos con valores muy discrepantes, a pesar de pertenecer a la misma especie animal.
Influencia del coeficiente de consanguinidad sobre el peso al nacimiento y peso al destete
Como antecedente contextual; Pond y Pond (2006) refieren que el peso al nacimiento, en explotaciones pecuarias, es importante para la sobrevivencia de la cría, está correlacionada con una posterior velocidad de desarrollo y al peso de venta. En llamas, es uno de los mejores indicadores tempranos de la producción de carne, que se mide a inicios de la vida del animal y constituye un buen predictor del comportamiento futuro (Apaza et al., 2016).
Al respecto, existen trabajos de investigación que determinan esta característica (Rodríguez y Martínez, 1976; Apaza y Pineda, 2001; Apaza, 2001; Bustinza y Gallegos, 1970; Sumar y Leyva, 1982; Paca y Bustinza, 1977; Málaga y Abarca, 1997; Frank y Freire, 1985). Lo evidente es que, de análisis de información de la data de referencia, este carácter no es muy variable, su valor es muy próximo a 10.60 kg en ambas variedades; valor medio que es ratificado por el resultado obtenido en el presente estudio (10.39±2.42 kg), enfatizando un mayor valor de la desviación estándar del mismo, que explica una mayor variabilidad de este carácter en este hato de llamas, el que se muestra ser diferente, altamente significante, por el efecto de la variedad y sexo.
Respecto al peso destete en llamas, de manera similar existen algunos reportes (Apaza, 2001; Apaza y Quispe, 1996; Ruiz y Mamani, 1991; Frank y Freire, 1985; Sumar y Leyva, 1982; Maquera, 1991; Pérez, 2008). Para el carácter peso al destete, los datos de referencia muestran gran variabilidad de la característica, diferencias atribuibles a distintas causas, como variedad, sexo, tamaño de la muestra utilizada para su determinación, el efecto de la edad al destete y su estandarización pertinente, el sistema de manejo y, finalmente la procedencia de los animales que representan a diferentes zonas agroecológicas. Los resultados obtenidos en el presente estudio de 38.62±6.02 kg ratifican este comportamiento diferente, además patentizan el efecto de las diferencias altamente significantes por el efecto de la variedad y sexo.
En este escenario es imprescindible cuestionarse sobre la magnitud con la que efecto genético de la consanguinidad contribuye el sobre la variabilidad de ambos caracteres en llamas, aspecto sobre el cual es exigua las referencias científicas.
La Tabla 5. presenta ecuaciones de regresión lineal del coeficiente de consanguinidad sobre el peso al nacimiento y peso al destete en llamas de las variedades K’ara y Ch’aku del Centro de Investigación y Producción La Raya de la UNA Puno, respectivamente.
Tabla 5 Ecuación de regresión y parámetros de ajuste para peso al nacimiento y destete en llamas

PENAC= peso al nacimiento, PDEST= peso al destete; C DE C = valor del índice del coeficiente de consanguinidad.
Un aspecto importante que se evidencia es el comportamiento del parámetro β (pendiente) en cada una de estas ecuaciones; en primera instancia, estos asumen un valor negativo para el peso al nacimiento (PENAC) en ambas variedades, y peso al destete (PDEST) de la variedad K'ara, situación que conlleva que a mayor valor del coeficiente de consanguinidad le corresponde menor peso, o que llamas con menores valores del coeficiente de consanguinidad tienen mayores pesos, en los caracteres referidos y en las variedades indicadas. De esta generalización es importante excluir al peso al destete de llamas de la variedad Ch'aku cuyo β es positivo, cuya inferencia sería contraria. Incongruentemente a lo manifestado, otro aspecto importante de resaltar es que los valores de este parámetro, para ambos caracteres y en ambas variedades de llamas, son muy pequeños e insignificantes (p) a la prueba del estadístico de T. Debilitando categóricamente la afirmación del comportamiento del coeficiente de consanguinidad respecto del peso vivo, lo que se traduce en una escaza influencia del efecto de la consanguinidad sobre ambos caracteres en esta especie animal.
La Tabla 6 complementariamente presenta la significancia (p) del efecto de la regresión en el estadístico de prueba F, para cada análisis de variancia de la regresión en ambas características y ambas variedades.
Tabla 6 Significancia de la fuente regresión para el peso al nacimiento, peso al destete en llamas.
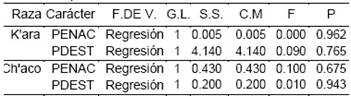
PENAC = peso al nacimiento; PDEST = peso al destete; F.DE V. = fuente de variabilidad; G.L.= grados de libertad; S.S. = suma de cuadrados; C.M. = cuadrado medio; F = estadístico de F; P = Significancia de F.
Para todos los casos, implica que la línea de regresión no es la función de ajuste eficiente de la distribución de datos, cuyo ajuste y explicación sobre las variables de interés son insignificantes, lo que se infiere de la interpretación de los valores de los coeficientes de determinación (R2) y determinación ajustados (R2 ajustado), comportamiento que puede deducir, además, por observación de los gráficos de las líneas de regresión en las Figuras 2 y 3, las que ratifican el comportamiento de dichos caracteres en ambas variedades de llamas.
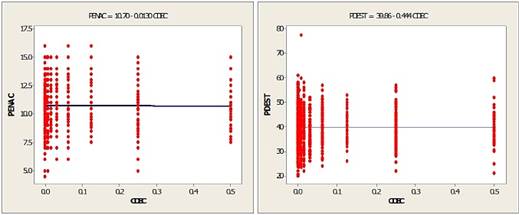
Figura 2 Regresión lineal del coeficiente de consanguinidad sobre el peso al nacimiento y destete en llamas de la variedad K'ara.
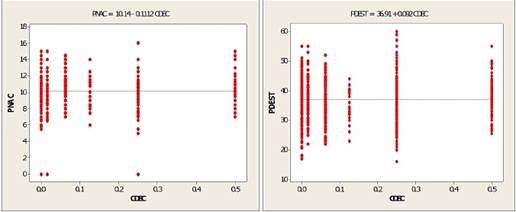
Figura 3 Regresión lineal del coeficiente de consanguinidad sobre el peso al nacimiento y destete en llamas de la variedad Ch'aku.
De estos resultados se colige que existe una relación inversa entre el coeficiente de consanguinidad y el peso al nacimiento y destete en llamas K'ara, así como en llamas Ch'aku, pero solo para el carácter peso al nacimiento; manifestándose una relación directa con respecto del peso al destete. Una interpretación genérica de este comportamiento implica que a un incremento promedio del valor de β le corresponde una disminución en una unidad promedio del valor del peso al nacimiento y/o destete. Para el caso al destete en llamas Ch'aku la relación directa implica que a un incremento promedio del valor β le corresponde un incremento promedio por cada unidad de cambio del peso al destete.
La Tabla 7, presenta valores el estadístico de F y la significancia para las fuentes de variación: año de producción (ADEP), variedad (variedad del animal), sexo (sexo del animal), edad de la madre (EDMA) y coeficiente de consanguinidad (C DE C), obtenidos de los Análisis de variancia para los caracteres peso al nacimiento y peso al destete en llamas.
De los resultados se infiere que todas las fuentes de variación tomadas en consideración, tienen efecto significante diferente en el peso al nacimiento y destete, excepto para el peso al destete en el que el factor coeficiente de consanguinidad no es significante en sus efectos, aspecto que puede interpretarse como que la depresión consanguínea, a pesar de que tiende un efecto significativo sobre el peso al nacimiento, por ser un carácter de escasa influencia ambiental externa, respecto del peso al destete, provoca su manifestación sobre caracteres que son más propensos a un efecto de la depresión consanguínea (Carrillo y Siewerdt, 2010; Davis y Simmen, 2010) y no así al destete, carácter que se ve ataviado por una serie de factores ambientales.
Tabla 7 Valores del estadístico F para las fuentes de variación: año de producción, variedad, sexo, edad de la madre y coeficiente de consanguinidad en el peso al nacimiento y destete en llamas.
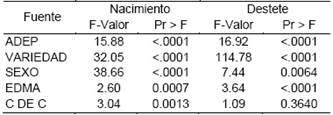
ADEP = año de producción; VARIEDAD = variedad de la llama; SEXO = sexo del animal; EDMA = edad de la madre; C DE C = coeficiente de consanguinidad del individuo.
CONCLUCIONES
El 40.13 % y el 34.00 % de las llamas K'ara y Ch'aku, respectivamente, no son consanguíneos, el 55.77 y 58.04 % de las llamas tienen coeficientes de consanguinidad muy bajas o bajas, entre 0.0015- 0.2500, de manera conjunta en ambas variedades; y el porcentaje remanente tienen un coeficiente de consanguinidad de 0.5000. La consanguinidad promedio poblacional para llamas de la variedad K'ara fue de 0.010 y la correspondiente a las de la variedad Ch'aku de 0.005, valores propios de estructuras genealógicas poblacionales poco profundas, generadas por un manejo ganadero carente de sistemas de apareamiento que permitan niveles de consanguinidad permisibles, y las características fisiológico reproductivos de esta especie animal que no conlleven a generar consanguinidad.
Los valores del parámetro β, para ambos caracteres y en ambas variedades de llamas, son muy pequeños e insignificantes (p) a la prueba del estadístico de T. La regresión lineal no genera una función de ajuste eficiente del coeficiente de consanguinidad individual sobre el peso al nacimiento y destete en llamas, precisa que no existe un efecto significativo de la consanguinidad sobre el peso al nacimiento y destete en llamas. Los coeficientes de consanguinidad individual no tienen influencia significativa sobre el peso al nacimiento y peso vivo al destete ajustado, en llamas de las variedades K'ara y Ch'aku.

